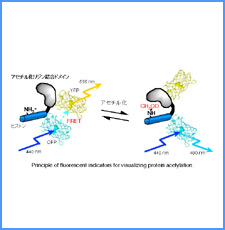
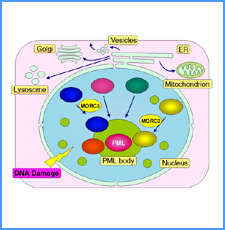
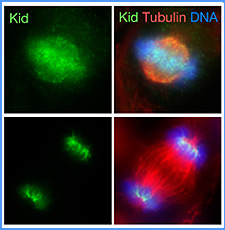
|
大山 隆
(早稲田大学教育・総合科学学術院・理学科生物学専修・教授)
研究分担者
三谷 匡
(近畿大学先端技術総合研究所・助教授)
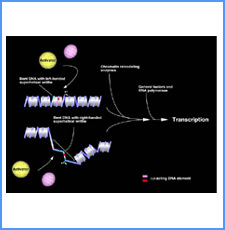
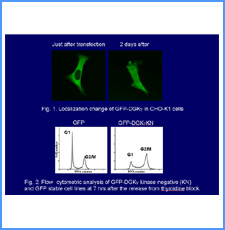
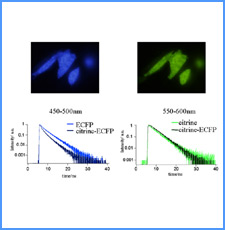
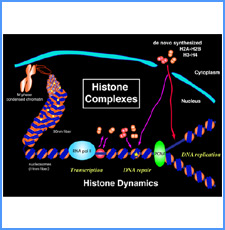
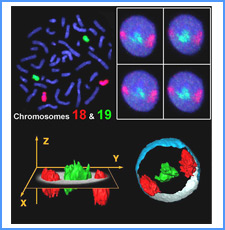
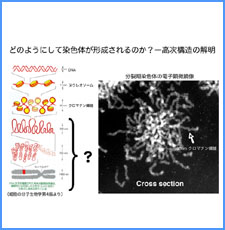
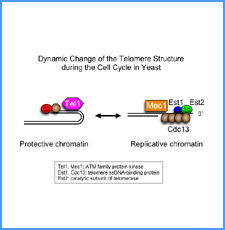
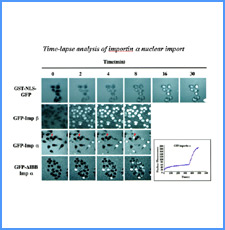
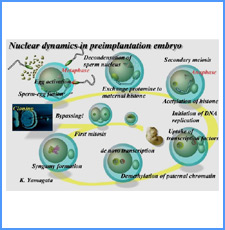
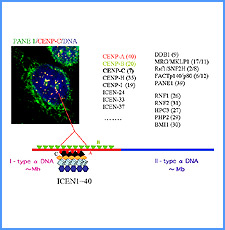
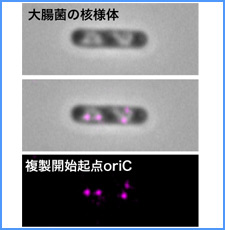
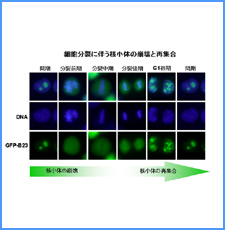
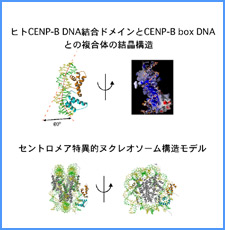
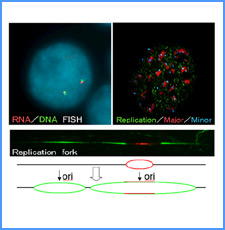
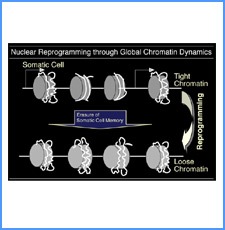
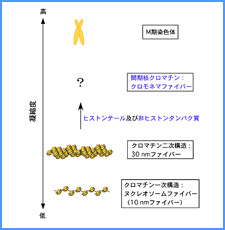
DNAのコンフォメーションや機械的特性がクロマチンの静態と動態にどのように関わっているかを明らかにする。我々はこれまでに、負の超らせんを擬態したベントDNAには、転写制御領域を転写開始可能な状態に保つ機能があることを見出した。また、クラスII遺伝子のプロモーターには、共通した特徴的な機械的特性があることを発見した。平成17年度は、マウスES細胞を用いて、ベントDNA構造のクロマチン制御能が細胞の分化に伴うクロマチンの静・動態の変動に影響されるかどうかを解析する。加えて、プロモーターに特徴的な機械的特性の生物学的意義について解析する。

|