自然科学研究機構 基礎生物学研究所
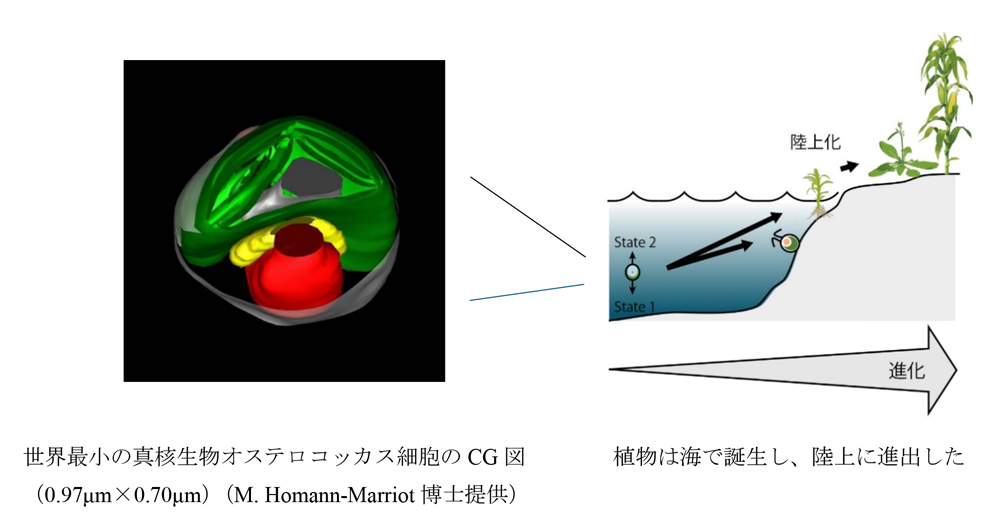
植物は光合成を通じて太陽光エネルギーを効率的に獲得しています。この過程で重要な役割を果たすのが、「ステート遷移」と呼ばれる光合成調節メカニズムです。ステート遷移は、青色光や赤外光などの特定波長の光を受けた際に、2つの光化学系(光化学系Iと光化学系II)の励起バランスを補正する仕組みです。緑色植物は大きく水生の「緑藻系統」と陸上の「ストレプト植物系統」に分類されます。これまでの研究から、どちらの系統においても、より進化した「陸上植物」や「コア緑藻」(クラミドモナスなど)がステート遷移を行っていることは知られていましたが、緑色植物すべてがこの能力を持つかどうかは不明でした。今回の画期的な発見は、緑藻系統とストレプト植物系統の共通祖先に近い海産の原始緑藻(プラシノ藻)もステート遷移を行うことが明らかになったことです。特筆すべきは、このステート遷移が従来知られていた陸上植物などのものとは異なり、青緑色光を利用していた点です。青緑色光は海中を満たしています。植物の祖先は海で誕生し、やがて陸上へ進出したと言われています。今回の研究結果は、ステート遷移の起源が植物がまだ海中に生息していた時代にまで遡ること、そして当初は海中での光合成調節メカニズムとして機能していたことを示唆しています。この発見により、生物進化と光化学系の進化の相互関係について新たな議論が喚起されると期待されます。本研究は、自然科学研究機構基礎生物学研究所の研究チーム(久保田真人大学院生、キム・ウンチュル助教、石井麻子研究員、皆川純教授)によって行われ、その成果は英国の専門誌New phytologist電子版(日本時間2024年9月24日付)に掲載されました。
【研究の背景】
太陽光は全ての生命にとって重要なエネルギー源です。光合成生物が集めた光は、光化学系I(PSI)と光化学系II(PSII)と呼ばれる二種類の装置により効率的にエネルギーに変換されています。効率の良いエネルギー変換を行うためには直列に配置されたこの2つの光化学系をバランス良く駆動する必要がありますが、太陽光の色や強さは刻一刻と変化するため、その励起バランスは崩れがちです。そのようなときにわずか数分でそのバランスを補正するしくみが「ステート遷移」です。緑色植物は大きく水生の「緑藻系統」と陸上の「ストレプト植物系統」に分類されますが、そのどちらにおいても、より進化した「コア緑藻」(クラミドモナスなど)や「陸上植物」では青色光や赤外光などの特定波長の光に対応して、ステート遷移が行われていることが知られています。しかし、全ての緑色植物がこの能力を持つかどうかは不明でした。特に緑藻と陸上植物が分岐した進化初期に誕生した種(原始緑藻)のPSIIとPSIは吸収スペクトルがよく似ていたため特定波長のもとでもバランスが崩れそうになく、これらの原始緑藻はステート遷移をしないと考えられてきました。その状況に疑問が生じたきっかけは、2023年に基礎生物学研究所を中心とするグループによって明らかとなったプラシノ藻オステロオコッカスの光化学系I(PSI)複合体の立体構造です。プラシノ藻は原始緑藻の一種であり、世界最小の真核生物としても知られる海産のピコ植物プランクトン
*1です。驚くべきことに、そのPSIには本来PSIIの集光アンテナであるはずの三量体Lhcp
*2がリン酸化されて結合していました(→
基礎生物学研究所プレスリリース2023年4月13日)。これまでの陸上植物や緑藻クラミドモナスを用いた研究から、ステート遷移はリン酸化された三量体LHCがPSII-PSI間を移動することがわかっていたため(→
基礎生物学研究所プレスリリース 2021年7月9日)、オステロコッカスがステート遷移を行っている可能性が出てきました。
【研究の成果】
研究チームは、最初にオステロコッカスのPSIとPSIIが持つ吸光特性を詳細に解析しました。その結果、陸上植物やコア緑藻と異なり、青緑色(480 nmから550 nm)やオレンジ色の光(650nm)の照射によってPSIIが選択的に励起され、PSIIとPSIの励起バランスを崩す可能性があることがわかりました (Fig.1)。大型スペクトログラフ
*3を利用してさまざまな波長の光を当ててみたところ、青緑光(480nm、520nm)やオレンジ光(650nm)の照射によリ実際に集光タンパク質Lhcpのリン酸化が見られ、またこの反応は時定数2分という速い反応であることがわかりました(Fig.2)。さらに、通常の植物は利用できない緑色光を照射する実験が行われました。その結果、緑色光によってリン酸化されたLhcpはPSIに結合しPSI-LHCI-Lhcp超複合体が形成されること(ステート2, Fig.3)、緑色光によるLhcpリン酸化はPSIIの阻害剤であるDCMUによって抑制されることなどがわかりました。これらの結果は、これまで陸上植物やコア緑藻で報告されてきたステート遷移と似た仕組みがオステロコッカスでも働いていることを意味しています。リン酸化されたLhcpは、細胞を暗黒下におくことで徐々に脱リン酸化されて元にもどる(ステート1)こともわかりました。これは、ステート遷移の重要な特徴である可逆的な反応特性を示しています。最後に、PSI-LHCI-Lhcp超複合体の形成によってPSIの集光能力が実際に増強されることを、細胞全体のクロロフィル蛍光寿命を測定することで確認し(Fig.4)、オステロコッカスはステート遷移を行うこと、そのためには特に青緑光の照射が効果的であると結論しました(Fig. 5)。
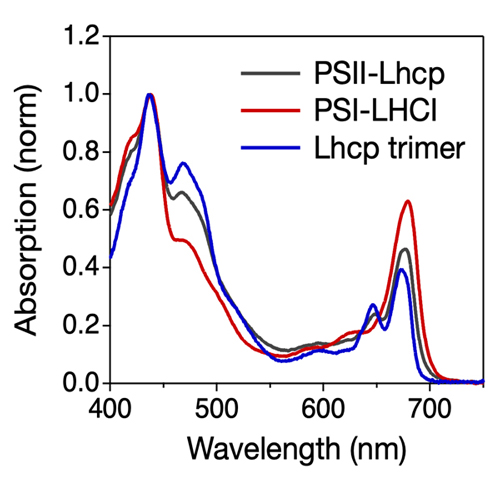 Fig.1. オステロコッカス光化学系の吸収スペクトル
オステロコッカス細胞からPSII-Lhcp超複合体 (灰色)、PSI-LHCI超複合体 (赤色)、Lhcp三量体 (青色) を単離し吸収スペクトルを測定した。PSII-LhcpやLhcpに特徴的な吸収帯が480nm, 520nm, 650nm付近にみられる。
Fig.1. オステロコッカス光化学系の吸収スペクトル
オステロコッカス細胞からPSII-Lhcp超複合体 (灰色)、PSI-LHCI超複合体 (赤色)、Lhcp三量体 (青色) を単離し吸収スペクトルを測定した。PSII-LhcpやLhcpに特徴的な吸収帯が480nm, 520nm, 650nm付近にみられる。
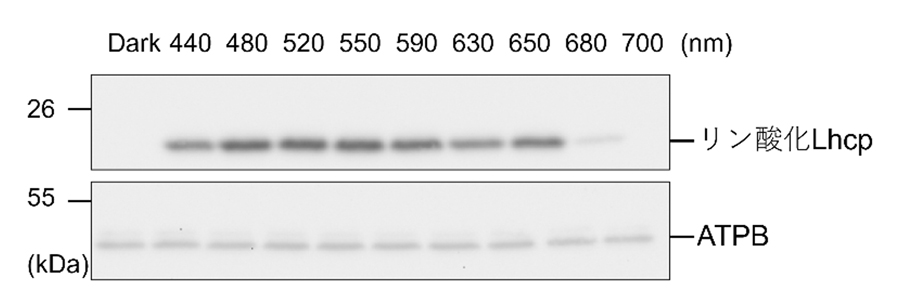 Fig.2. Lhcpリン酸化を誘導する単色光の調査
抗原抗体反応を用いて特定タンパク質を検出するウエスタンプロット解析によりLhcpのリン酸化が誘導される光環境を調べた。青緑色である480 nm から550 nm の照射によるリン酸化の強度が強いことがわかる。
Fig.2. Lhcpリン酸化を誘導する単色光の調査
抗原抗体反応を用いて特定タンパク質を検出するウエスタンプロット解析によりLhcpのリン酸化が誘導される光環境を調べた。青緑色である480 nm から550 nm の照射によるリン酸化の強度が強いことがわかる。
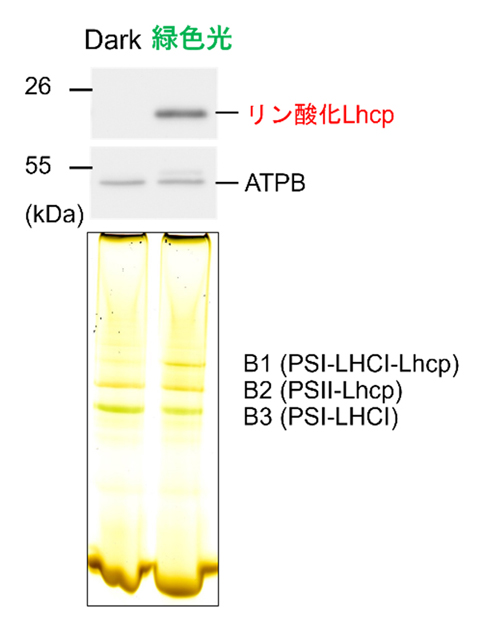 Fig.3. 緑色光照射によるPSI-LHCI-Lhcp超複合体の形成
Fig.3. 緑色光照射によるPSI-LHCI-Lhcp超複合体の形成
Clear Native-PANGE(CN-PAGE)と呼ばれる電気泳動により光合成超複合体を分離した。緑色光を10分間照射してLhcpをリン酸化すると、PSI-LHCI超複合体(B3)にさらにLhcpが結合したPSI-LHCI-Lhcp超複合体(B1)が出現した。
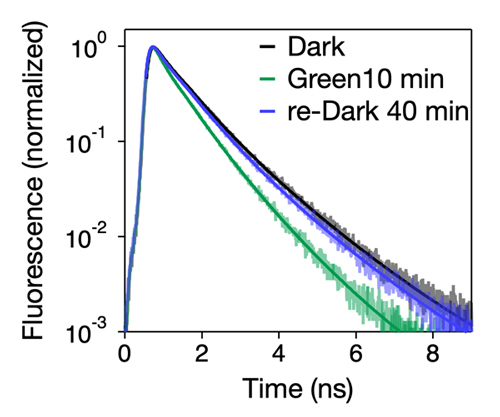 Fig.4. 緑色光照射によるクロロフィル蛍光寿命への影響
Fig.4. 緑色光照射によるクロロフィル蛍光寿命への影響
クロロフィル結合タンパク質のクロロフィルを励起すると、一般に赤色のクロロフィル蛍光が見られる。PSIIとPSIはどちらもクロロフィル蛍光を発するが、PSIIからの蛍光と比べ、PSIからの蛍光ははるかに寿命が短い。この原理を利用して、どちらの光化学系がより光を集めているかを調べることができる。ここでは、緑色光10分間照射により全体の蛍光寿命が短縮され、暗黒条件に戻すことで寿命が長くなることが示されている。緑色光照射でステート遷移(ステート1→ステート2)がおこりPSI-LHCI-Lhcp超複合体が形成された結果、PSIの集光能力が拡大したことがわかる。
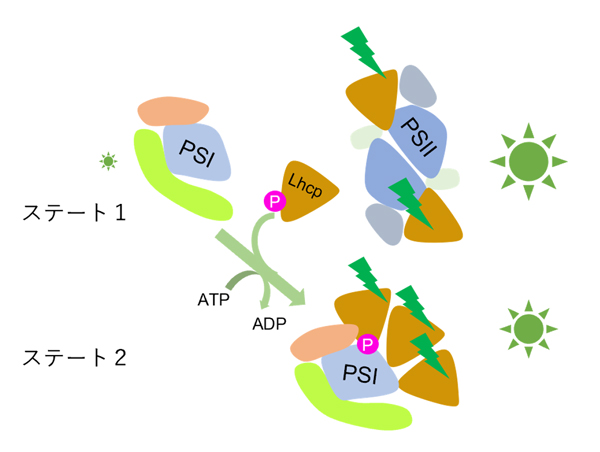 Fig.5. オステロコッカスにおけるステート遷移モデル
暗黒条件では、LhcpはPSIIに結合しているためPSIの集光能力は低い(ステート1)。青緑光などの照射によりLhcpが選択的に励起されるとLhcpはリン酸化を受けてPSIIから脱離しPSIに再結合する。この結果、PSI-LHCI-Lhcp超複合体が形成されて、PSIの集光能力が増強される(ステート2)。時定数2分という短い時間でおこるこの現象は、遺伝子発現調節によらず集光能力を調節する仕組み(ステート遷移)として知られ、原始緑藻の時代から植物が備えていた能力であることが明らかとなった。
【本研究の意義と今後の展開】
Fig.5. オステロコッカスにおけるステート遷移モデル
暗黒条件では、LhcpはPSIIに結合しているためPSIの集光能力は低い(ステート1)。青緑光などの照射によりLhcpが選択的に励起されるとLhcpはリン酸化を受けてPSIIから脱離しPSIに再結合する。この結果、PSI-LHCI-Lhcp超複合体が形成されて、PSIの集光能力が増強される(ステート2)。時定数2分という短い時間でおこるこの現象は、遺伝子発現調節によらず集光能力を調節する仕組み(ステート遷移)として知られ、原始緑藻の時代から植物が備えていた能力であることが明らかとなった。
【本研究の意義と今後の展開】
今回の発見は、ステート遷移(ステート1⇔ステート2)が緑色植物の誕生初期から備わっていた重要な光合成最適化機構であることを示しています。特に注目すべきは、ここで見られたステート遷移が従来知られていた青色光/赤外光によってではなく、青緑色光/暗黒によって誘導されるものであったことです。オステロコッカスなどの原始的な藻類が海産の植物プランクトンであること、水中に届く光が主に青や緑色であることを考え合わせると、この「青緑色型ステート遷移」は、海中での光合成に必要であると考えられます。原始緑藻がこのような「青緑色型ステート遷移」を行っていたということから、以下のような推論が成り立ちます。植物がまだ植物プランクトンとして紫外線を避けて海の中で生活をしていた太古の昔、ほとんど光の届かない深い層ではステート1の状態にありますが、鉛直撹拌が起こって上層に浮かび上がった際には青や緑色の光を浴びてステート2に移行したのかもしれません。今後の研究で、海の上層でステート2になりPSIの集光能力を強化することの意義が明らかなることが望まれます。さらに研究が進むと、植物がやがて陸上化して太陽光のフルスペクトルを浴びるようになり、それにしたがってステート遷移も青色光/赤外光への適応へと変化した理由も明らかにされることでしょう。生物の進化に光合成系の進化がどのように影響を与えてきたのか、視野が広がります。
【用語解説】
*1ピコ植物プランクトン:プランクトン(水生の浮遊生物)のうち、体長0.2~2µmで光合成を行うもの。あまりに小さいため旧来のプランクトンネットでは採集できず、最近になって研究が進んだ。
*2 Lhcp:真核光合成生物は一般にクロロフィルやカロテノイドを多数結合した集光タンパク質を用いて光合成に必要な太陽光を集める。LHC(Light-Harvesting Complex)は陸上植物などが有する集光タンパク質だが、Lhcpはプラシノ藻類特有の集光タンパク質である。広い意味ではどちらも同じ系統のタンパク質群に属する。Lhcpはプラシノキサンチンなどの480nmから540nm(青色~緑色)に吸収帯を持つカロテノイドを多数結合していることが特徴として知られる。
*3 大型スペクトログラフ:基礎生物学研究所地下に設置されている世界最大の分光照射装置。30kWのキセノンアークランプから照射される光は赤道直下の太陽光と同程度の強度を持ち、これを多波長に分光したものが生物実験に用いられる。大学共同利用機関として世界中から研究者が利用に訪れている。
【掲載誌情報】
雑誌名:
New phytologist
論文タイトル: The blue-green light-dependent state transition in the marine phytoplankton
Ostreococcus tauri
著者: Masato Kubota , Eunchul Kim , Asako Ishii, and Jun Minagawa
DOI:
https://doi.org/10.1111/nph.20137
URL:
https://nph.onlinelibrary.wiley.com/doi/10.1111/nph.20137
【研究サポート】
本研究は科学研究費補助金(学術変革領域研究「光合成ユビキティ」(23H04960)、基盤研究(S) (21H05040))、基礎生物学研究所共同利用研究 (24NIBB605)のサポートを受けて行われました。
【問い合わせ先】
基礎生物学研究所 環境光生物学研究部門
教授 皆川 純(ミナガワ ジュン)
〒444-8585 愛知県岡崎市明大寺町字西郷中38
TEL: 0564-55-7515
E-mail: minagawa@nibb.ac.jp
ホームページ:
https://www.nibb.ac.jp/photo/
【報道担当】
基礎生物学研究所 広報室
TEL: 0564-55-7628
FAX: 0564-55-7597
E-mail: press@nibb.ac.jp




