京都府立大学 生命環境科学研究科
自然科学研究機構 基礎生物学研究所
金沢大学 ナノ生命科学研究所
熊本大学 大学院先端科学研究部
植物の根毛は、細長い形状を維持しながら伸長します。これは、先端が伸長すると同時に、根毛側面部分は、二次細胞壁が形成されることで、その膨張が抑制されるためです。しかしながら、根毛側面に二次細胞壁成分を輸送する分子機構については、全く明らかになっていませんでした。この度、京都府立大学の平野朋子准教授、佐藤雅彦教授らを中心とした共同研究グループ
*は、モデル植物シロイヌナズナを用いて、
根毛の側面部分に二次細胞壁成分を輸送し、根毛側面を硬くすることで、根毛が細長く真っ直ぐ伸びながら、その形を維持する仕組みを解明しました。
今後、この仕組みを活用して、側面強度を増強した長い根毛を持つ植物体を作出するなど、
栄養源が乏しい土壌中から効率よく栄養を吸収できる植物体を開発できる可能性があります。
本研究成果は、
国際学術誌 “The Plant Cell”に 2023年9月15日にオンライン掲載されました。
論文タイトル:The SYP123-VAMP727 SNARE complex is involved in the delivery of secondary cell wall components for hardening the root hair shank in Arabidopsis
著者:Tomoko Hirano, Kazuo Ebine, Takashi Ueda, Takumi Higaki, Takahiro Watanabe-Nakayama, Hiroki Konno, Hisako Takigawa-Imamura, and Masa H. Sato
doi.org/10.1093/plcell/koad240
【研究概要】
植物の根の表皮細胞は、「根毛」を形成する細胞(根毛細胞)と形成しない細胞(非根毛細胞)が交互に配列しています。「根毛」は、根毛細胞の一部が突き出て管状に伸びた構造で、根の表面積を大きくして土壌中の水や養分を吸収する役割があります。
根毛細胞が均等に伸長した場合、風船状に膨らみますが(図1)、
実際の根毛は、先端部分が伸びると同時に根毛の側面部分の拡大を抑制するために、細長い管状構造を形成しています。また、
土の抵抗に逆らって破けず細長く伸びるために、側面を非常に硬くしています。このような硬い根毛側面には、
二次細胞壁が形成されていますが、二次細胞壁成分を輸送する分子機構については、全く不明でした。
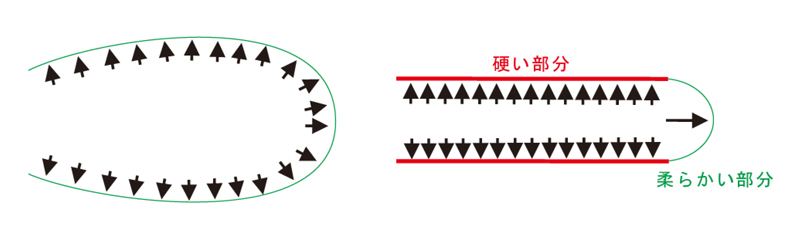 図1
図1.根毛は、内部からの膨圧のはたらきなどにより伸長することから、根毛の細胞壁の硬さが一様な場合、風船状に膨らんでしまい、細長い構造を維持できない。根毛は、側面に二次細胞壁成分が輸送されることで、硬く細長い真っ直ぐな構造が作られる。
今回、京都府立大学を中心とする共同研究グループは、モデル植物シロイヌナズナを用いて、根毛の「先端成長のための物質輸送ルート」と「側面の硬化と成長抑制のための物質輸送ルート」が存在することを発見しました。さらに、これらのルートの担い手は、前者が、SYP132,VAMP721の複合体と、SYP123,VAMP721の複合体で、後者が、SYP123とVAMP727の複合体であることも突き止めました(図2)。
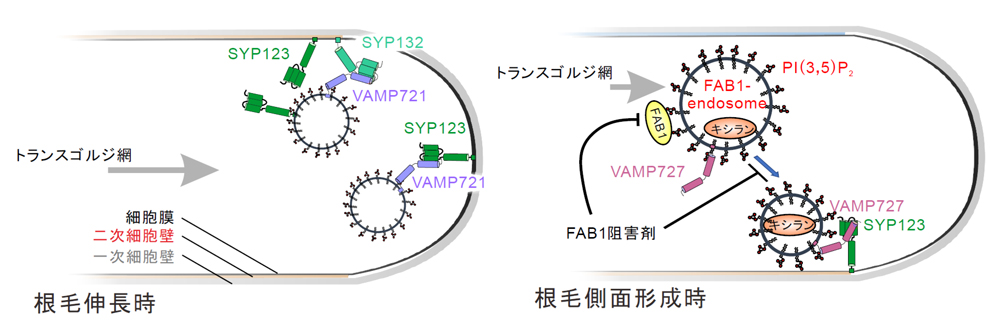 図2
図2. 研究成果の概略図
伸長中の根毛では、SYP123とVAMP721の複合体や、SYP132とVAMP721の複合体が働く「先端成長のための物質輸送ルート」とSYP123とVAMP727の複合体が働く「側面の硬化と成長抑制のための物質輸送ルート」が存在する。
*共同研究グループ
・京都府立大学大学院 生命環境科学研究科 (研究統括、実施全般)
准教授 平野 朋子
教授 佐藤 雅彦
・自然科学研究機構基礎生物学研究所 細胞動態研究部門(全反射顕微鏡解析)
助教 海老根 一生
教授 上田 貴志
・金沢大学 ナノ生命科学研究所(原子間力顕微鏡解析)
准教授 紺野 宏記
准教授 中山 隆宏
・熊本大学 大学院先端科学研究部(微小管解析)
教授 檜垣 匠
・九州大学大学院 医学研究院系統解剖学分野(数理生物学的解析)
助教 今村 寿子
【本件に関するお問い合わせ】
<研究内容に関するお問い合わせ>
京都府立大学 生命環境科学研究科 細胞動態学研究室 教授 佐藤雅彦
TEL/FAX:075-703-5448 E-mail:mhsato@kpu.ac.jp
<報道関係のお問い合わせ>
京都府立大学 企画・地域連携課
TEL:075-703-5212 E-mail:kikaku@kpu.ac.jp
自然科学研究機構 基礎生物学研究所 広報室
TEL:0564-55-7628 E-mail:press@nibb.ac.jp
金沢大学 ナノ生命科学研究所 広報・事業企画G
TEL:076-234-4550 E-mail:nanolsi-office@adm.kanazawa-u.ac.jp
熊本大学総務部総務課 広報戦略室
TEL:096-342-3271 E-mail:sos-koho@jimu.kumamoto-u.ac.jp
【研究の詳細】
1.研究の背景
土壌で細長く伸びる根毛は、硬くて、真っ直ぐな構造である。
植物の根毛は、土壌中から水分や養分を効率よく吸収するために発達した細長い構造体で、根の表皮細胞の一部が細長く突出して形成されます。
著者らの先行研究により、根毛の側面形成には、イノシトールリン脂質
注1の一種であるホスファチジルイノシトール3,5-二リン酸[PI(3,5)P
2]
注2とその合成酵素FAB1と低分子量GTPase ROP10が関与することがわかっていました。すなわち、これら分子の複合体形成による「根毛側面の細胞膜直下の表層微小管の構造の安定化」と、 PI(3,5)P
2を目印とした「二次細胞壁成分の輸送と分泌」を、PI(3,5)P
2が制御し、細長く、真っ直ぐな、側面が硬化した根毛構造が形成されるということです(図3)。しかしながら、PI(3,5)P
2が制御する二次細胞壁の輸送と分泌がどのように行われているかについては、ほとんどわかっていませんでした。
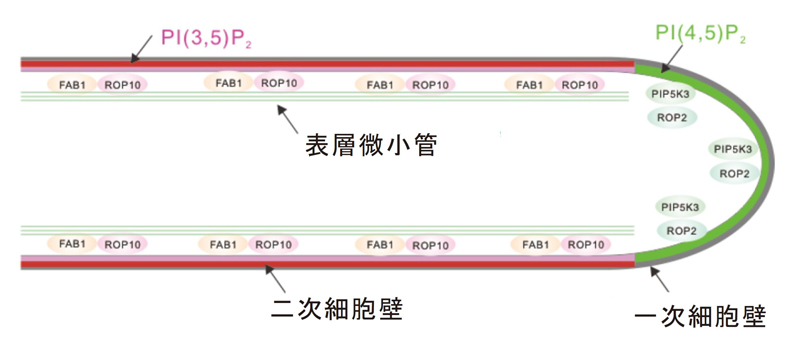 図3
図3.二種類のイノシトールリン脂質のうち、PI(4,5)P
2は先端成長に働き、その異性体PI(3,5)P
2は根毛側面の硬化に働く。
SYPタンパク質とVAMPタンパク質が物質の輸送場所を決定する。
一方で、著者らは、植物を含む真核生物の細胞内において、膜で囲まれた細胞小器官(オルガネラ)間の物質輸送システムである「小胞輸送」の研究より、SYP132、SYP123というSYPタンパク質とVAMP721、VAMP727というVAMPタンパク質が根毛の伸長に関与することを明らかにしていました。
「小胞輸送」は、「出発地のオルガネラの膜がくびれて、輸送される分子を包んで出芽して輸送小胞が生成され、これが目的地のオルガネラ膜(標的膜)に融合」して、小胞体やゴルジ体で合成された分子を輸送するシステムです。輸送小胞に存在するVAMPタンパク質ファミリーと標的膜に存在するSYPタンパク質ファミリーの組み合わせで、融合オルガネラの特異性、すなわち、物質輸送場所の特異性が決められます(図4)。
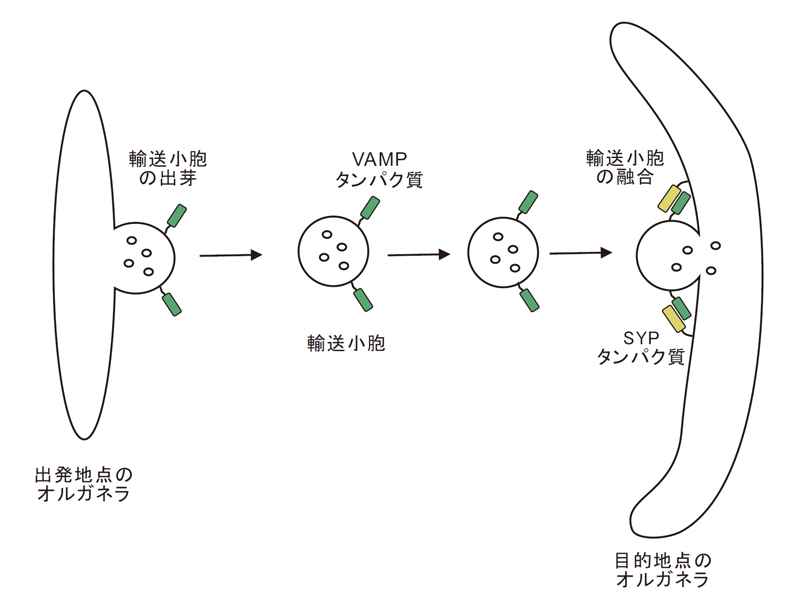 図4
図4.真核生物の細胞内小胞輸送は、「出発地のオルガネラの膜がくびれて、輸送される分子を包んで出芽して輸送小胞が生成され、これが目的地のオルガネラ膜(標的膜)に融合」して輸送するシステム
2.本研究の成果
SYP123は、FAB1/PI(3,5)P
2が制御する輸送経路で働き、二次細胞壁成分を輸送する
根毛の細胞膜に特異的に発現するSYP123と緑色蛍光タンパク質(GFP)を融合したタンパク質 (GFP-SYP123)を発現する形質転換シロイヌナズナの根毛を観察すると、GFP-SYP123は、伸長開始と伸長停止時は、細胞膜全域に均等に配置していましたが、伸長中では、根毛先端よりも側面に強く局在していました(図5)。
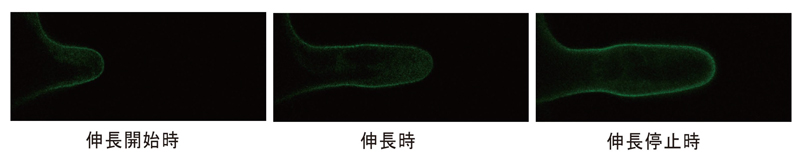 図5
図5.シロイヌナズナ根毛では、GFP-SYP123は、伸長開始時と伸長停止時に細胞膜全体に、伸長時に側面細胞膜に局在する。
そして、GFP-SYP123は、PI(3,5)P
2の生成を阻害すると、細胞内に顆粒状に蓄積し、細胞膜に局在できなくなりました。また、SYP123を欠失したシロイヌナズナ(
syp123変異体)は、根毛が太くて短い形態を示し、これは野生型でPI(3,5)P
2の生成を阻害した場合と同様でした(図6)。
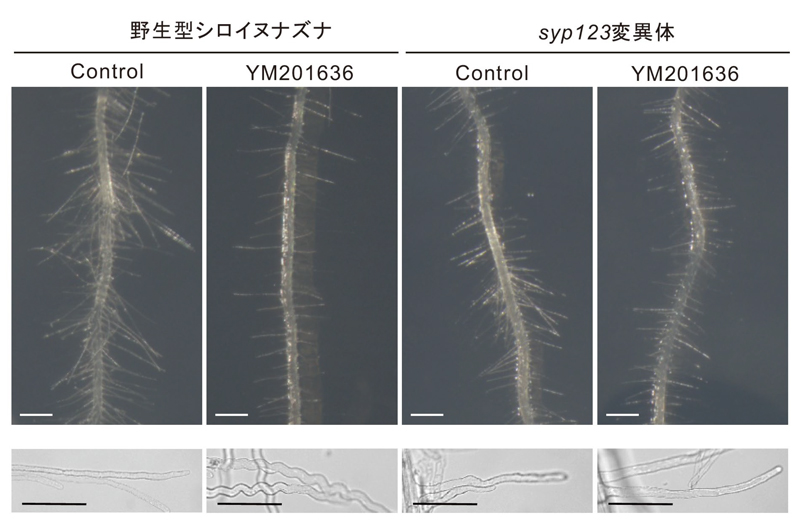 図6.PI(3,5)P2合成酵素、FAB1阻害剤YM201636を処理した野生型とsyp123変異体の根毛の形態変化
図6.PI(3,5)P2合成酵素、FAB1阻害剤YM201636を処理した野生型とsyp123変異体の根毛の形態変化
以上のことから、FAB1およびPI(3,5)P
2が制御する輸送経路と、SYP123が働く輸送経路は、同一の経路であることが示唆されました。
また、著者らは先行研究において、原子間力顕微鏡
注3を用いた根毛側面強度の直接的測定法により、PI(3,5)P
2の生成を抑制した植物の根毛側面強度が極度に低下していることを示していました。
そこで、
syp123変異体の根毛側面強度を測定したところ、やはり著しい低下が観察されました(図7)。これは、
syp123変異体の根毛で、二次細胞壁の成分であるキシランとリグニンの蓄積が低下しているためであることが明らかになりました。
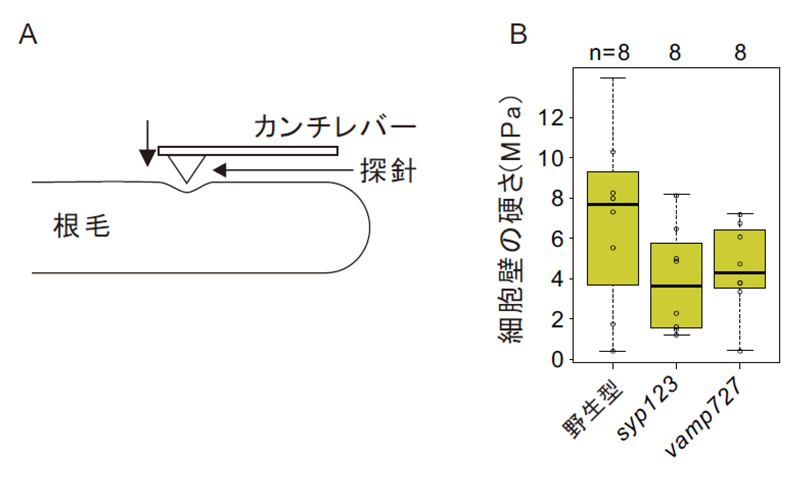 図7.syp123とvamp727変異体は、根毛側面の硬さが減少している
図7.syp123とvamp727変異体は、根毛側面の硬さが減少している
(A)原子間力顕微鏡は、探針を根毛側面に押し付け、カンチレバーのたわみを測定することで、試料の硬さを測定することができる。(B)
syp123や
vamp727変異体の根毛側面は、野生型の半分の硬さである。
SYP123は、VAP727とペアで二次細胞壁成分を輸送する
一方、二分子蛍光相補 (BiFC)法
注4と共免疫沈降法
注5により、SYP123は、輸送小胞に存在するVAMP721とVAMP727の両方と相互作用することを証明しました(図8)。さらに、SYP123とVAMP727の相互作用は、根毛側面の細胞膜上で、二次細胞壁成分を分泌する際に生じることを、共焦点レーザー顕微鏡
注6と全反射顕微鏡
注7を用いて示しました(図9)。
これらの結果により、SYP123は、VAMP721が関与する輸送経路とVAMP727が関与する輸送経路の二種類の輸送経路に関与していることが明らかとなりました。
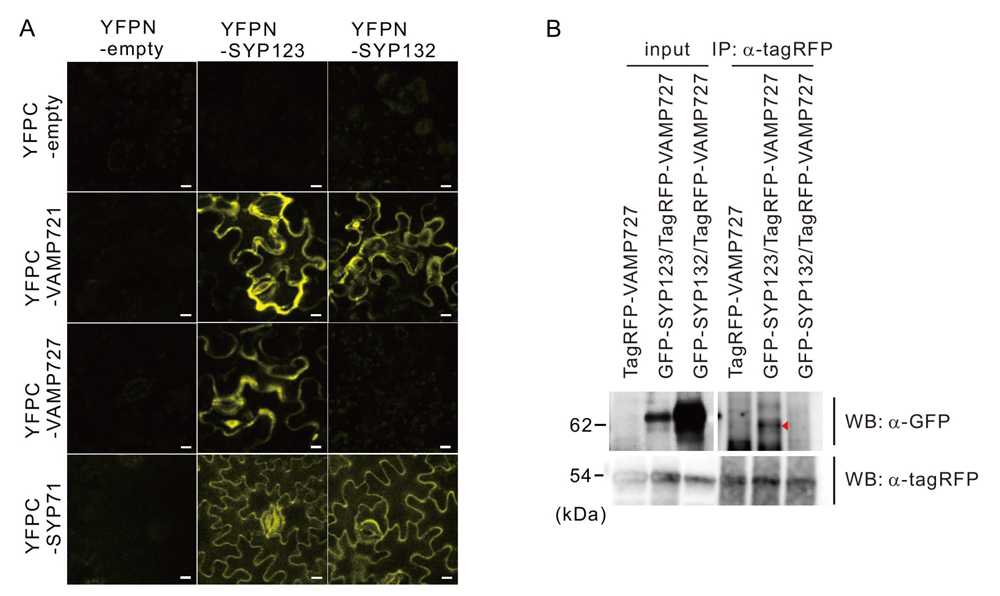 図8.SYP123は、VAMP727とVAMP721の両者と相互作用する
図8.SYP123は、VAMP727とVAMP721の両者と相互作用する
(A)二分子蛍光相補(BiFC)法による相互作用解析。SYP123は、VAMP727とVAMP721の両者と相互作用するが、SYP132は、VAMP721のみと相互作用する。(B)共免疫沈降法による相互作用解析。SYP123はVAMP727と相互作用する。
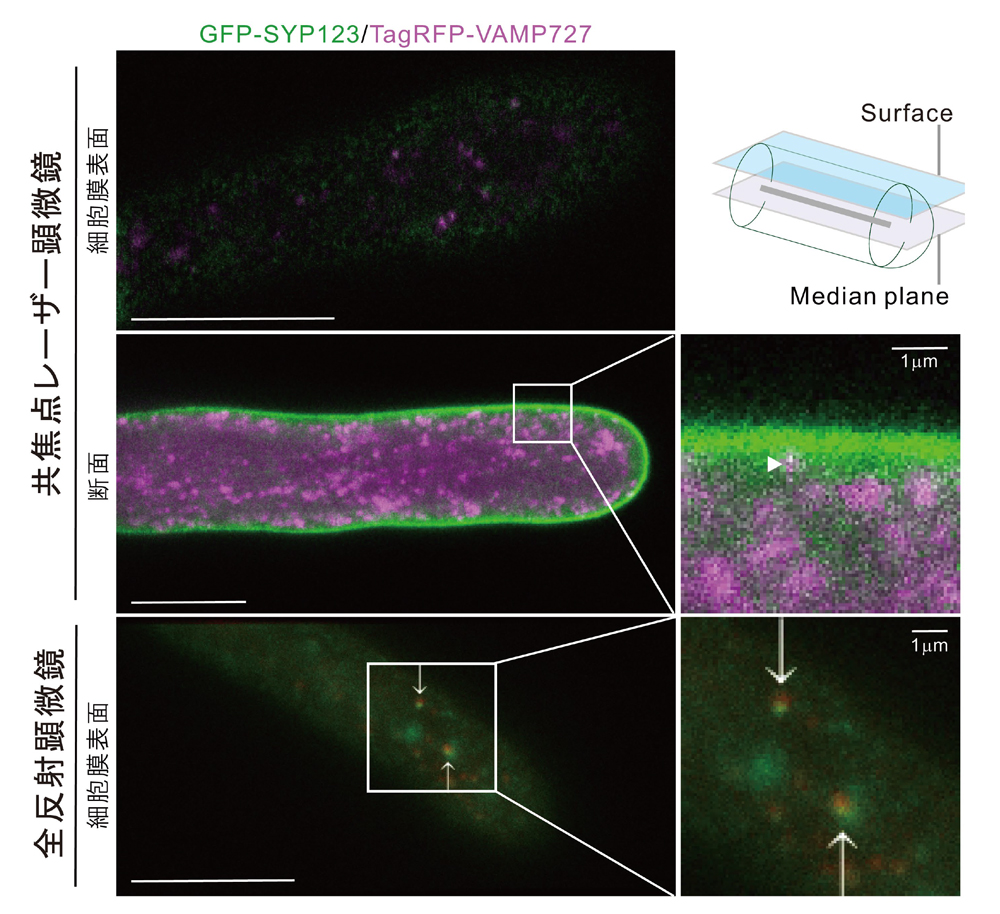 図9.共焦点レーザー顕微鏡と全反射顕微鏡により、GFP-SYP123とTagRFP-VAMP727は、細胞膜周辺で共局在が観察された。
図9.共焦点レーザー顕微鏡と全反射顕微鏡により、GFP-SYP123とTagRFP-VAMP727は、細胞膜周辺で共局在が観察された。
表層微小管は根毛の細長い真っ直ぐな形態をつくる
著者らは以前に、FAB1/PI(3,5)P
2の機能阻害が、根毛側面の硬さの低下と細胞膜直下の表層微小管
注8の断片化を引き起こし、根毛が太くて短く、規則的に波打つ形態を示すことを報告していました。しかし本研究では
syp123変異体の根毛は、太くて短いものの、波打つ形態は観察されませんでした。そこで、FAB1/PI(3,5)P
2の機能阻害植物と
syp123変異体の根毛側面の硬さと表層微小管の安定性をパラメーター化し、力学的な計算機シミュレーションを行い、根毛の形態において、二次細胞壁と表層微小管のそれぞれの寄与を切り分けることに成功しました(図10)。
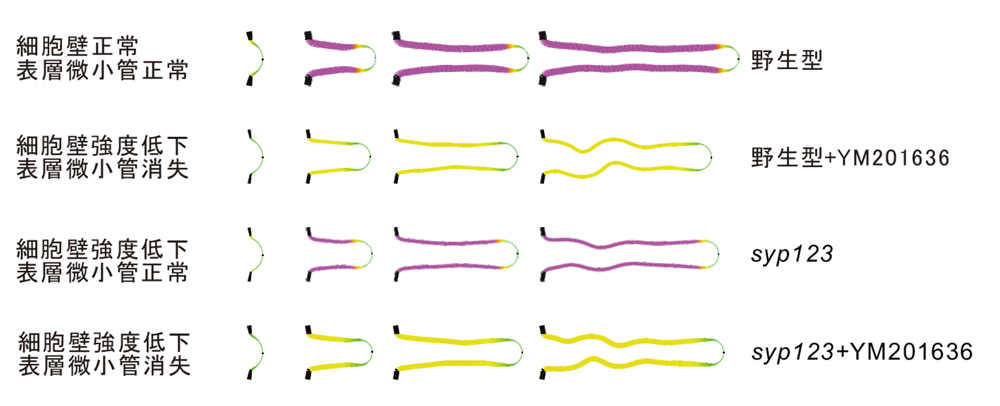 図10.力学的な計算機シミュレーション
図10.力学的な計算機シミュレーション 根毛では、表層微小管が形態を決め、二次細胞壁の輸送・分泌が形態維持する役割があり、これらの事象は独立している。
以上の結果より、根毛細胞では、「先端成長のための物質輸送ルート」にSYP132とVAMP721の複合体とSYP123とVAMP721の複合体が、「側面の硬化と成長抑制のための物質輸送ルート」に、SYP123とVAMP727の複合体が働くことで、根毛を伸長すると同時に側面を硬くし、根毛の細長い形を作っていることが明らかとなりました。
・用語説明
注1:イノシトールリン脂質
イノシトールリン脂質は、リン脂質のリン酸基部分にイノシトールが結合したもの。生体膜に存在するリン脂質総量の約1割程度と微量で存在し、生体膜上のシグナル分子やランドマークとして、様々な細胞機能をに関わる。
注2:ホスファチジルイノシトール3, 5-二リン酸 [PI(3,5)P
2]
イノシトールリン脂質のイノシトール環の3位と5位の水酸基がリン酸化されたものが、ホスファチジルイノシトール3,5-二リン酸 [PI(3,5)P
2]であり、ホスファチジルイノシトール-3リン酸 5-キナーゼ、であるFAB1により、ホスファチジルイノシトール3-リン酸(PI3P)の5位の水酸基がリン酸化されることで生成する。
注3:原子間力顕微鏡 [Atomic Force Microscopy (AFM)]
片持ち梁(カンチレバー)の先端にある鋭く尖った探針と試料の間に働く相互作用力をカンチレバーで検出する走査プローブ顕微鏡の一種であり、試料表面を走査することにより、表面形状と粘弾性を直接測定できる。
注4:二分子蛍光相補 (BiFC;Bimolecular Fluorescence Complementation)法
二つに分割した蛍光タンパク質(黄色蛍光タンパク質(YFP)など)のそれぞれの断片に、相互作用を調べたいタンパク質をそれぞれ融合し、これらのタンパク質を同時に発現させて蛍光の有無でタンパク質間の相互作用を検出する方法。目的の2つのタンパク質同士が、物理的に相互作用すると、蛍光タンパク質断片同士が近接して再構築し、蛍光を発することを利用している。
注5:共免疫沈降法
試料溶液中に存在するタンパク質複合体を、タンパク質複合体に含まれるタンパク質の抗体を用いて、捕捉・精製する方法。精製に用いた抗体と別の抗体を用いて、相互作用するタンパク質を検出することができる。
注6:共焦点レーザー顕微鏡
蛍光顕微鏡の一種であるが、レーザー光を用いて励起した試料の特定の焦点面のみの蛍光像を検出するため、厚みのある試料でもピントをあわせた画像を得ることが可能である。
注7:全反射顕微鏡
蛍光顕微鏡の一種である対物レンズの中心から外れた位置から励起光を斜めに入射させ、スライドガラスと試料の境界面で励起光をほとんどすべて反射させ(全反射)、スライドガラスとの境界面に発生するエバネッセント光で試料を励起し、観察する。エバネッセント光は、スライドガラスの境界面から数百ナノメートルのみにしか発生しないため、細胞膜上や、細胞膜近傍のタンパク質の挙動の観察等に用いられる。
注8:表層微小管
微小管は、αチューブリンとβチューブリンが構成単位となって形成される中空の繊維状構造で、細胞膜直下に存在する微小管のことを表層微小管と呼ぶ。表層微小管には、方向性をもった配向により、細胞壁の主成分であるセルロース微繊維の方向を制御し、細胞の伸長方向を決定する役割がある。
・発表雑誌 The Plant Cell doi.org/10.1093/plcell/koad240
論文タイトル:The SYP123-VAMP727 SNARE complex is involved in the delivery of secondary cell wall components for hardening the root hair shank in Arabidopsis
著者:Tomoko Hirano, Kazuo Ebine, Takashi Ueda, Takumi Higaki, Takahiro Watanabe-Nakayama, Hiroki Konno, Hisako Takigawa-Imamura, and Masa H. Sato
・研究サポート 本研究は科学研究費補助金、基盤研究A (19H00933)、基盤研究B(16H06280)、基盤研究C (20K05962)、住友財団などの支援により実施されました。




