自然科学研究機構 基礎生物学研究所
マメ科植物は、窒素固定細菌(根粒菌)を細胞内に取り込むことでコブ状の共生器官「根粒」を形成します。この現象は根粒共生と呼ばれ、マメ科植物はこの共生により大気中の窒素を栄養素として効率よく利用できます。今回、基礎生物学研究所の後藤崇支 大学院生(総合研究大学院大学、日本学術振興会特別研究員DC2)、征矢野敬 准教授、森友子 技術班長、川口正代司 教授らは、根粒共生の成立過程において、植物ホルモンの1種であるオーキシン(インドール-3-酢酸:IAA)がメチル化されることが、共生成立に重要であることを発見しました。
これまでに、オーキシンのメチル化酵素であるIAMT1はシロイヌナズナ(非マメ科植物)の葉や茎の成長に関わることが知られていましたが、根での機能は不明でした。本研究では、表皮感染を介した根粒共生を進化の過程で獲得したマメ科植物においてIAMT1が遺伝子重複により2つ存在していること、そして、その片方の遺伝子(
IAMT1aと命名)が根粒菌感染によってミヤコグサ
(注1)の根の表皮で誘導されることを発見しました。
IAMT1aの機能をRNAi法により抑制すると、直下の皮層で起こる根粒形成が抑制されました。これにより、オーキシンのメチル化が、根粒の形成に必要とされることが明らかとなりました。
また、オーキシン (IAA) のメチル化産物であるMeIAAをミヤコグサの根に与えると、根粒共生の鍵遺伝子である
NINの発現が誘導されました。この結果は、共生におけるMeIAAの重要性を示すものです。シロイヌナズナ等で得られた知見から、これまでオーキシンのメチル化は、オーキシンの機能を不活性化するものであると考えられてきました。しかし本研究により、根粒共生の成立過程において、オーキシンメチル化は単なる不活性化とは異なる機能を持っていることが示されました。
本研究成果は、2022年3月2日付けで米国科学アカデミー紀要(Proceedings of the National Academy of Sciences of the United States of America)誌に掲載されました。
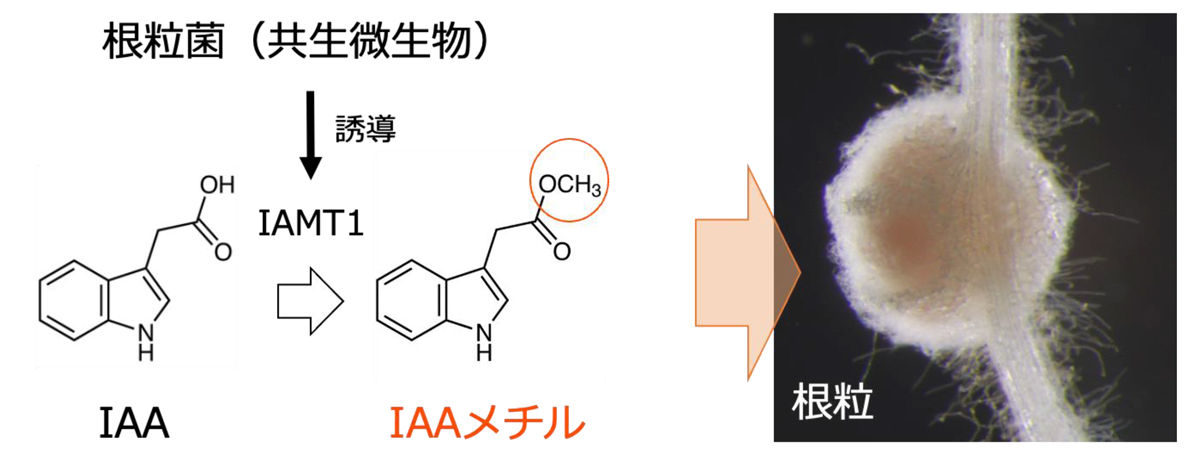
図1 オーキシンのメチル化
オーキシンメチル化酵素(IAMT1)によって、オーキシン(IAA)はIAAメチル(MeIAA)へと変換(メチルエステル化)されます。本研究は、根粒菌の感染によって誘導されるオーキシンのメチル化が、根粒の形成に重要であることを明らかにしました。
注1)ミヤコグサ:マメ科植物の遺伝子機能を探るためのモデル植物。
【研究の背景】
マメ科植物は、根の表皮から共生細菌(根粒菌)を取り込むことで、「根粒」と呼ばれる窒素固定器官を形成します。根粒の形成は、表皮における根粒菌の感染過程と、皮層における細胞分裂を伴う原基の形成過程から構成されます(図2)。皮層における細胞分裂は感染した表皮の直下で起こり、根粒菌が表皮から感染糸を介して分裂する皮層細胞に取り込まれることが、根粒の形成に必要不可欠です。
これまでに、感染した表皮で生成された未知のシグナルが、皮層へと送られ、皮層細胞分裂を誘導することが示唆されてきましたが、その分子実体は長らく不明でした。
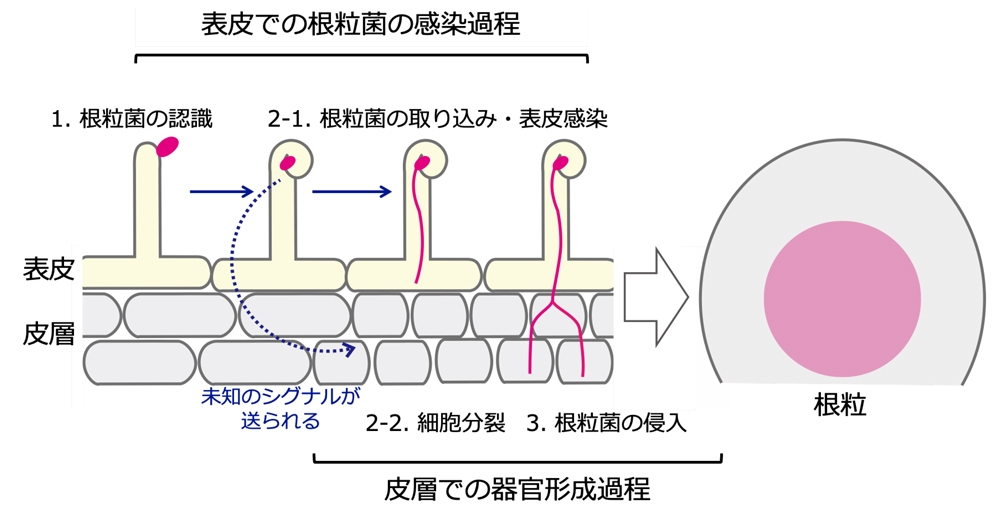
図2 根粒ができるまでの過程
表皮と皮層の共生イベントは連動して進行します。皮層での細胞分裂は、表皮での根粒菌感染時に生じた未知のシグナル(青色)によって誘導されることが示唆されていますが、その分子実体は長らく不明でした。
【研究の成果】
基礎生物学研究所の後藤崇支 大学院生と川口正代司 教授らは、マメ科のモデル植物であるミヤコグサの共生変異体
daphne(表皮での根粒菌感染は過剰に生じるが、皮層での共生イベントが進行しない変異体:図3)
(注2)と正常な野生型との間で、共生初期過程に発現する遺伝子について、時系列RNA-seqによる網羅的な比較解析を行いました。その結果、変異体
daphneと野生型との間で異なる発現パターンを示した遺伝子の1つとして
IAMT1(オーキシンメチル化酵素遺伝子)が見いだされました。
IAMT1は植物ホルモンとして知られるオーキシンをメチル化する酵素です(図1)。これまでにシロイヌナズナ(非マメ科植物)での解析により、葉や茎の成長に関わることが知られていますが、根における関与は不明でした。
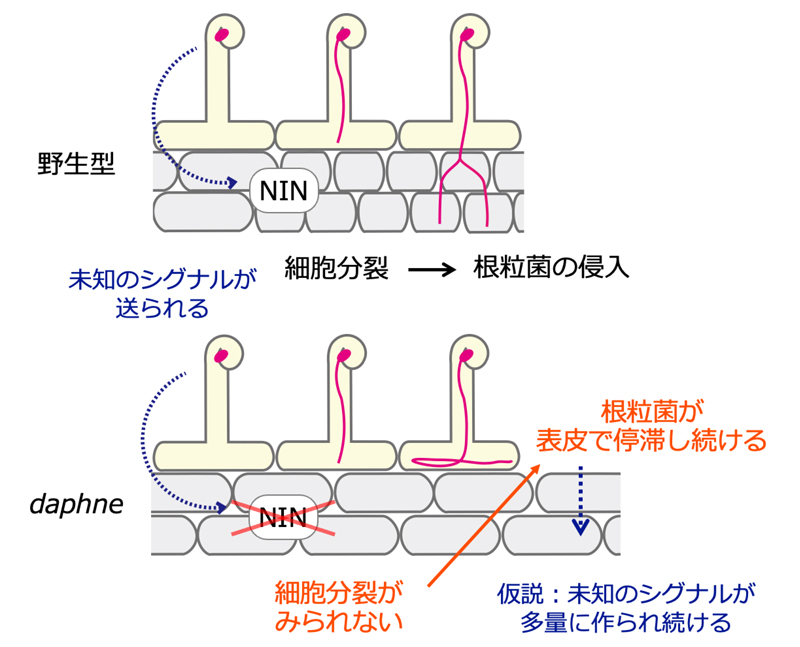
図3 野生型と変異体
daphneの共生初期過程
daphneは皮層細胞分裂の誘導に関わる“表皮由来”の未知のシグナルを同定するための有効な変異体です。皮層細胞分裂の鍵遺伝子
NINのプロモーター領域を欠損した
daphneでは、皮層での細胞分裂が起きません。一方で表皮では根粒菌が広範囲に感染しますが、皮層細胞分裂が起きないために根粒菌は表皮で停滞します
(注2)。ゆえに表皮から皮層へと送られる未知のシグナルは
daphneでは多量に作られていると考えられます。
注2)養老、川口らによる過去の研究報告
https://www.nibb.ac.jp/pressroom/news/2014/05/30.html
ミヤコグサの根において、根粒菌感染によって
IAMT1の発現が誘導されることが分かったため、LC-MS/MS解析を通して、オーキシンのメチル化産物であるMeIAAが、実際に共生の現場で作られるか調べました。そして根粒菌の過剰な感染が見られる
daphneの根でMeIAAが野生型より多く蓄積していることを検出することに成功しました(図4)。
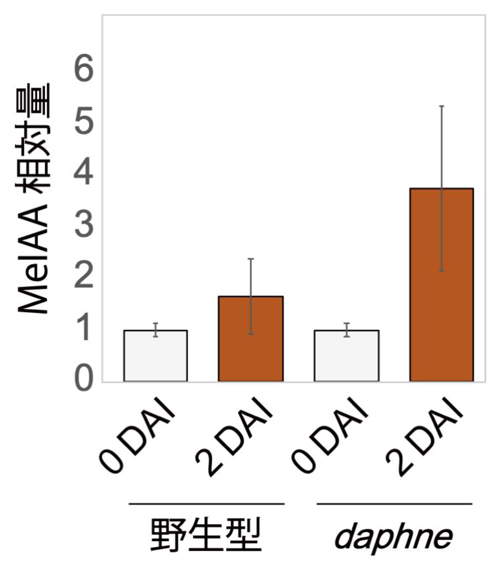
図4 根粒菌感染前後(感染前:DAI0 → 感染2日後:DAI2)におけるMeIAA量の変化
様々な植物の
IAMT1のアミノ酸配列を比較し分子系統樹を作成した結果、
IAMT1はミヤコグサ、大豆、ピーナッツ、インゲン豆、 小豆などで遺伝子が2つずつ見つかり、マメ科植物の系統で遺伝子重複していることが示唆されました(図5)。
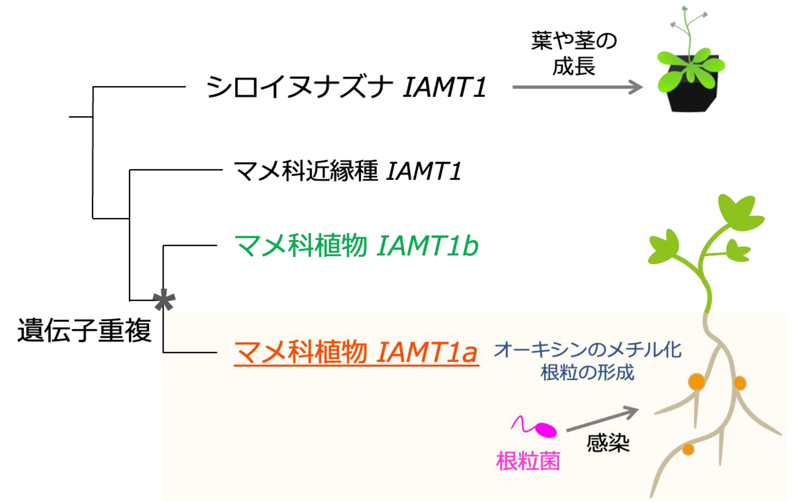
図5 マメ科植物の系統におけるIAMT1の遺伝子重複
2つのIAMT1遺伝子のうち、根粒菌感染によって誘導されるIAMT1をIAMT1aと命名。
研究グループは、
IAMT1aが根粒菌感染によって根の表皮で誘導されることを発見しました。RNAi法を用いて
IAMT1aの発現を抑制すると、根粒形成が強く抑制され、
daphneと同様に根粒菌が皮層へと侵入できず表皮で停滞している様子が観察されました(図6)。
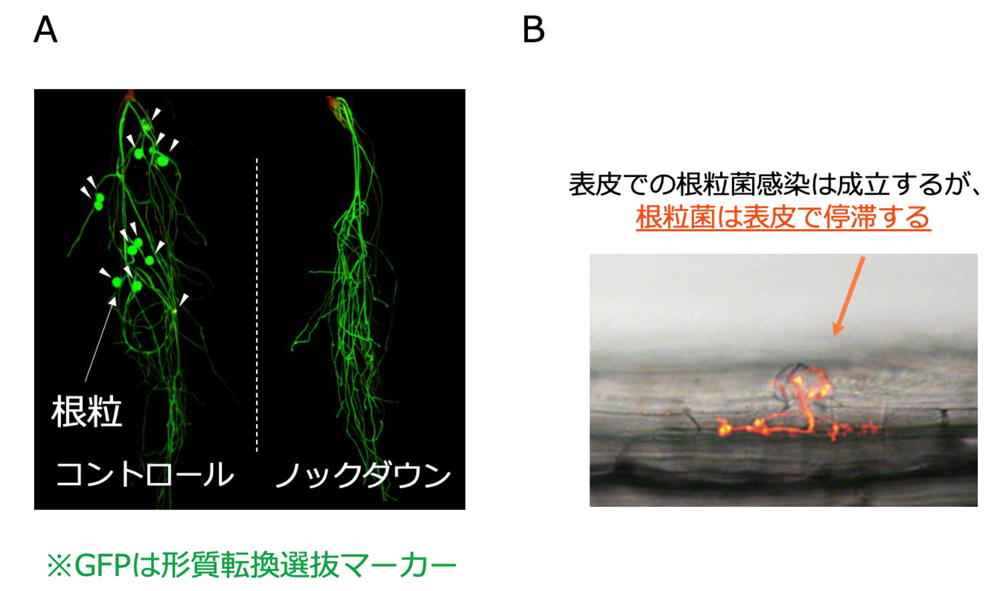
図6
IAMT1aをRNAi法によりノックダウンすると、根粒(矢頭)の形成が抑制されました(A)。
IAMT1aをノックダウンした根では、根粒菌の表皮感染の成立には影響を受けませんが、根粒菌が表皮で停滞していることが確認されました(B)。
興味深いのは、
IAMT1aは根粒菌感染に伴い根の表皮で強く発現誘導されるものの、その機能は表皮での感染成立ではなく皮層での共生イベントに必要とされる点です。これまで、シロイヌナズナでの解析から、オーキシンのメチル化は、細胞内でオーキシン量を調整する(減らす)“不活化プロセス”であると考えられてきましたが、根粒共生の成立過程において、MeIAAには表皮から皮層へと組織を超えて働きかける未知の分子機能があるのかもしれません。
この可能性を支持するように、MeIAAをミヤコグサの根に投与すると、根粒菌を感染させていないにも関わらず根粒共生の鍵遺伝子であり皮層細胞分裂を誘導する
NIN遺伝子の発現が顕著に誘導されました(図7)。これはMeIAAがIAAとは異なる分子機能を有していることを示唆しています。
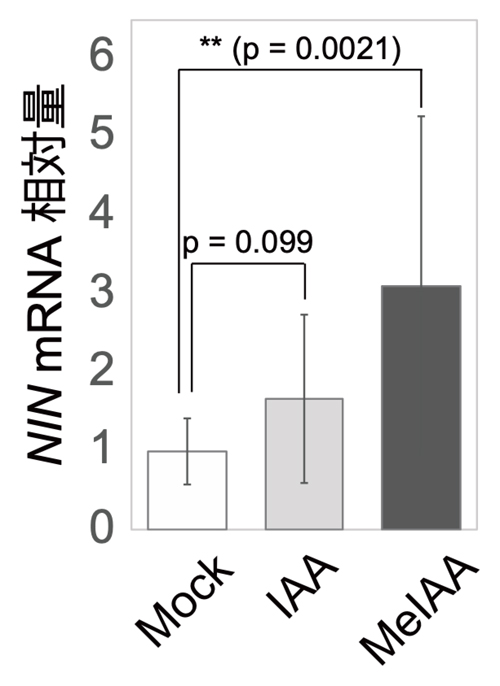
図7:メチル化オーキシン(MeIAA)添加によるNIN遺伝子発現の誘導
【まとめと今後の展望】
本研究により、表皮での感染と皮層での発生という異なる現象をつなぐ新たな共生シグナリングと分子が見えてきました(図8)。また、表皮感染を介した根粒共生を進化の過程で獲得したマメ科植物における
IAMT1の遺伝子重複そして
IAMT1aの表皮での発現は、マメ科植物に保存された根粒共生システムの頑強性に貢献しているかもしれません。今後
IAMT1の遺伝子重複の起源を探るとともに、MeIAAを起点としたシグナリングを解明することで、根粒共生成立の分子機構と進化の解明が進むと思われます。また本研究は、これまで一括りに不活化プロセスと見なされていたオーキシンのニ次代謝に新たな光を当てる成果です。
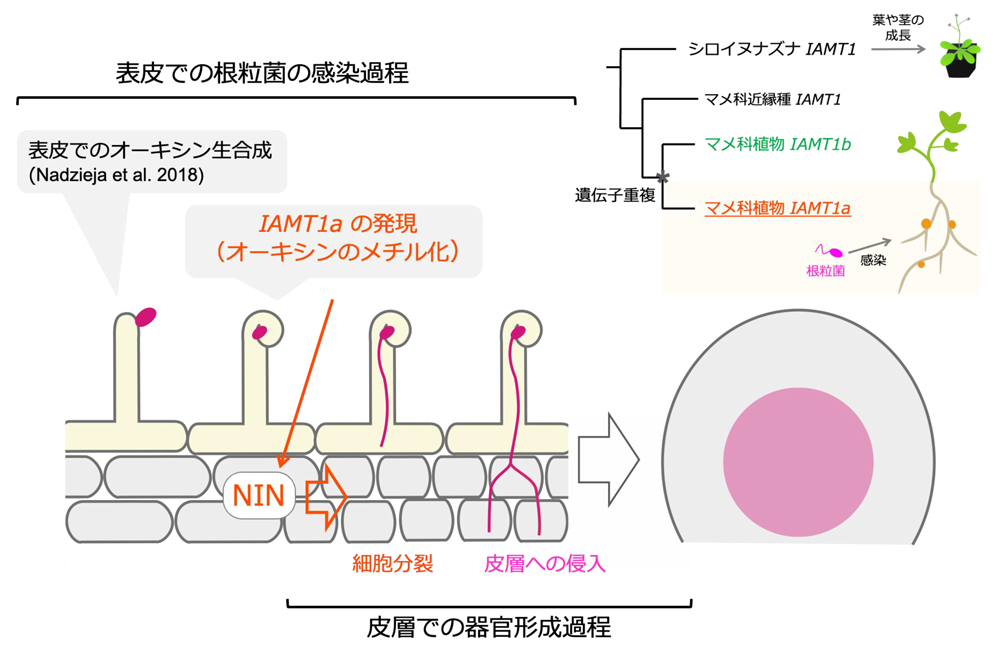
図8 本研究のまとめ:マメ科植物の系統における遺伝子重複で生じた
IAMT1aは、根粒菌の感染によって根の表皮で発現が誘導されます。IAMT1aはIAAをMeIAAに変換し、皮層で細胞分裂の鍵遺伝子
NINの発現の誘導に関わります。
【発表雑誌】
雑誌名 米国科学アカデミー紀要Proceedings of the National Academy of Sciences of the United States of America (PNAS)
掲載日:2022年3月2日
論文タイトル:Auxin methylation by
IAMT1, duplicated in the legume lineage, promotes root nodule development in
Lotus japonicus
著者:Takashi Goto*, Takashi Soyano, Meng Liu, Tomoko Mori, Masayoshi Kawaguchi.
(*: co-corresponding author)
DOI:
https://doi.org/10.1073/pnas.2116549119
【研究グループ】
本研究は、基礎生物学研究所 共生システム研究部門と、同研究所 生物機能情報分析室によって実施されました。
【研究サポート】
本研究は、科学研究費助成事業 (21J10143、20H03283、19K05959) と基礎生物学研究所の生物機能情報分析室、情報管理解析室、光学解析室のサポートを受けて実施されました。
【本研究に関するお問い合わせ先】
基礎生物学研究所 共生システム研究部門
大学院生・日本学術振興会特別研究員DC2 後藤崇支
TEL: 0564-55-7563
E-mail: tgoto@nibb.ac.jp
教授 川口正代司
TEL: 0564-55-7564
E-mail: masayosi@nibb.ac.jp
【報道担当】
基礎生物学研究所 広報室
TEL: 0564-55-7628
FAX: 0564-55-7597
E-mail: press@nibb.ac.jp




