基礎生物学研究所



2014.05.30
基礎生物学研究所共生システム研究部門の大学院生養老瑛美子(日本学術振興会特別研究員)、寿崎拓哉助教、川口正代司教授らの研究グループは、これまで根粒共生の正の制御因子として知られていたNODULE INCEPTION (NIN)遺伝子が、根粒菌感染の過程を負に制御しているという新たな機能的側面を発見しました。本研究は、根粒共生の成立過程で異なる組織で起こる複数の現象間での相互作用の一端を明らかにしたものです。本研究成果は、植物学の専門誌Plant Physiology誌6月号に掲載されます(電子速報版は4月10日付で公開されました)。
【研究の背景】
マメ科植物と根粒菌の共生は、植物の根に形成されるこぶ状の器官である根粒を介して成立しています。この共生関係の成立過程は、「感染」と「器官発生」に大別されます。根粒形成における「感染」とは、根の最外組織である表皮への根粒菌の付着、根毛のカーリング、感染糸*1の形成と伸長、という一連の過程を指します。一方、「器官発生」は、より内側の組織である皮層において、細胞が分裂して原基を形成することを指します。この皮層由来の新たに分裂した細胞に、表皮から感染糸を介して侵入した根粒菌が内部共生し、根粒が形成されます。すなわち、「感染」と「器官発生」の二つ現象が、表皮と皮層という異なる組織で同調的に進行することが根粒形成にとって重要です(図1)。
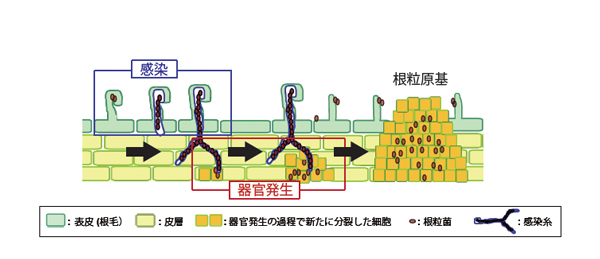
図1. 根粒共生の主なイベント:「感染」と「器官発生」
根粒共生は、表皮で起こる「感染」と、皮層で起こる「器官発生」に大別される。
これまでに、マメ科のモデル植物ミヤコグサを中心として、根粒共生に異常を示す変異体が単離されてきました。しかし、その多くの変異体は「感染」と「器官発生」の両者に欠陥をもつため、この二つの現象の同調的進行の過程における表皮と皮層の組織間でのシグナル伝達については、ほぼ未解明でした。
また、NODULE INCEPTION (NIN)遺伝子はRWP-RKドメインをもつ転写因子をコードしており、変異により根粒が形成されず、さらにNIN遺伝子を過剰に発現させると、根粒菌非存在下でも根粒様の構造が形成されることから、根粒共生において正の役割を果たすと考えられてきました。
【本研究の成果】
本研究では、はじめにイオンビームを照射したミヤコグサ種子から、新たな共生の表現型を示す変異体を単離しました。daphne(ダフネ)*2と名付けたこの変異体は、根粒は全く形成しないが、感染糸の数が野生型に比べて20倍増加する過剰感染糸形成という珍しい表現型を示しました(図2)。daphne変異体の原因遺伝子座を同定したところ、既に根粒共生の正の制御因子として知られているNIN遺伝子の上流に染色体転座が生じており、daphne変異体原因遺伝子はNINであること示唆されました。ただし、NIN遺伝子の完全機能欠失変異体ninは、根粒のみならず感染糸をも形成できないため、感染糸を形成できるdaphneとは異なります。
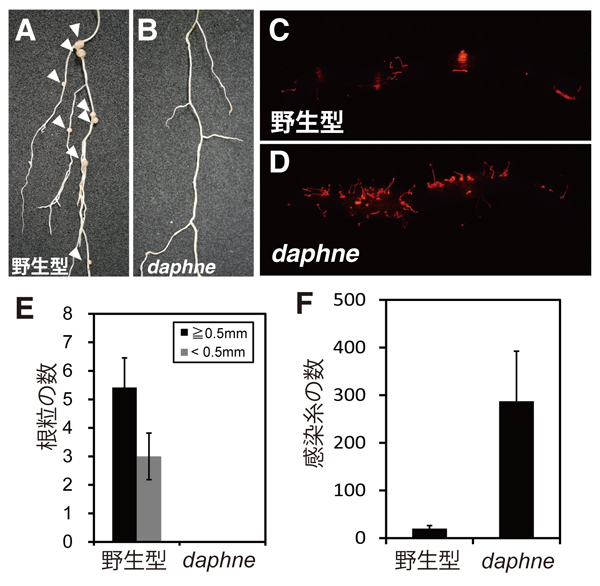
図2. daphne変異体の表現型
daphne変異体では、根粒は全く形成されないが(A,B,E)、感染糸は過剰に形成される(C,D,F)。
そこで、NIN遺伝子の発現パターンを解析した結果、daphne変異体では根粒菌感染によって誘導されるNIN遺伝子の根全体の発現量は低下しているが、表皮における発現領域は拡大していました。また、daphne変異体においてNIN遺伝子を過剰に発現させると、過剰感染糸形成は強く抑制されました(図3)。
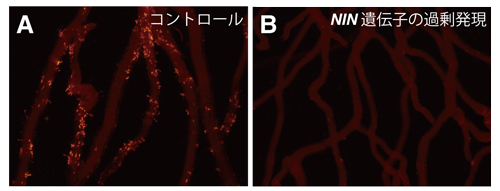
図3. daphne変異体におけるNIN遺伝子の過剰発現による感染糸形成抑制効果
コントロールとしてGUS遺伝子を過剰発現させた根では過剰感染糸形成のままであるが(A)、NIN遺伝子を過剰発現させた根では感染糸形成は強く抑制された(B)。
さらに、「器官発生」が起こる皮層の組織特異的エンハンサー(J0571)*3のもとで、NIN遺伝子を発現させると、過剰感染糸形成の抑制のみならず、根粒形成の復帰が観察されました。これらの結果から、daphne変異体は、NIN遺伝子の機能のうち、根粒菌感染初期の「感染」における正の機能(図4 青線で囲ったNIN)は維持しているものの、後期の「器官発生」に由来して「感染」を負に制御する機能(図4 赤線で囲ったNIN)が失われている変異体であることが明らかになりました。
本成果により、私たちはマメ科植物がNIN遺伝子を介して根粒菌の感染量を調節するメカニズムについての新たなモデルを提唱しました。まず、根粒菌感染後初期(図4-(1))において、表皮(根毛を含む)で発現するNIN遺伝子は「感染」を正に制御します(図4 青線で囲ったNIN)。一方、根粒菌感染後期(図4-(2)-(3))へと進むにつれ、皮層で発現するNIN遺伝子が「器官発生」を正に制御し細胞分裂を開始すると同時に、表皮における「感染」を負に制御し(図4 赤線で囲ったNIN)、過剰な根粒菌感染を妨げているのではないかというモデルです。daphne変異体では、初期のNIN遺伝子の機能を維持しつつ、後期のNIN遺伝子機能が失われたために、過剰な根粒菌感染が生じたと考えられます(図4 daphne変異体)。さらに、その過剰な根粒菌感染は、皮層でNIN遺伝子を発現させることで、後期のNIN遺伝子機能を回復させることで、感染量は減少した(図4 daphneでJ0571エンハンサーでNIN遺伝子を発現)といえます。
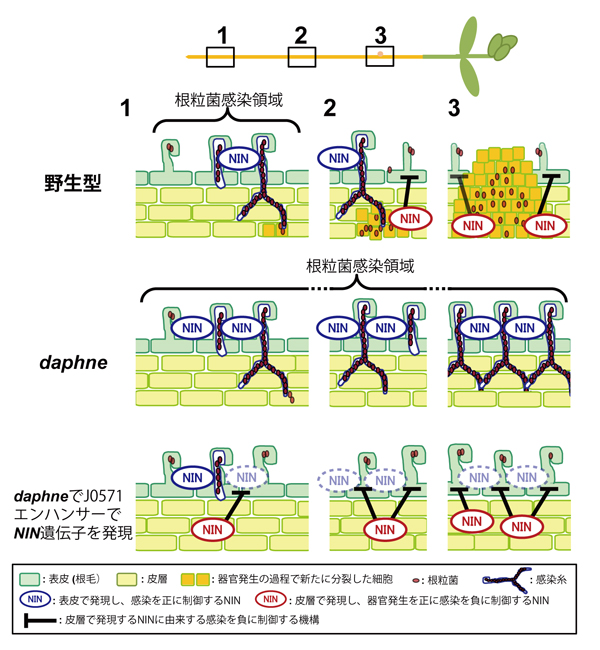
図4.NIN遺伝子を介して「感染」を負に制御するモデル
根粒共生の進行段階を1-3で示す。根粒菌感染初期においては、表皮で発現するNINが「感染」を正に制御し(青線で囲んだNIN)、後期において皮層で発現するNINは「器官発生」を正に制御すると同時に、「感染」を負に制御している(赤線で囲んだNIN)。
これまでに、私たちの研究グループは根粒の数(つまり「器官発生」の過程)を負に制御する因子として、CLE-RSペプチド、HAR1、 KLV、 TMLを同定し、その制御メカニズムについて報告してきました。しかし、根粒菌の「感染」を制御する仕組みについては、ほとんどが未解明です。今後、このNIN遺伝子のもつ「感染」における負の機能的側面は、根粒菌感染過程の制御機構の解明と、その生物学的意義の理解について進展をもたらすことが期待されます。
【補足説明】
*1感染糸
マメ科植物が、共生菌である根粒菌を根組織内部へと導くために形成するトンネル状の構造体で、植物の細胞膜が陥入することで形成される。本研究では、根粒菌に赤色蛍光タンパク質を発現させ、根粒菌を可視化することで感染糸を観察している。
*2 daphne(ダフネ)
アポローンからの求愛を拒み続けるギリシャ神話の女神の名前に由来。根粒菌は過剰に感染するが、根粒を形成せず、根粒共生は成立しないという表現型から名付けた。
*3皮層の組織特異的エンハンサー(J0571)
シロイヌナズナの根の組織のうち、内皮と皮層で特異的に遺伝子発現を誘導するシス配列として同定された。本研究では、NIN遺伝子の表皮で起こる「感染」と皮層で起こる「器官発生」の機能を区別するため、NIN遺伝子の「器官発生」の機能のみを誘導させる目的で用いた。
[論文情報]
Plant Physiology (プラント・フィジオロジー)
著者:Emiko Yoro, Takuya Suzaki, Koichi Toyokura, Hikota Miyazawa, Hidehiro Fukaki, and Masayoshi Kawaguchi
[研究グループ]
本研究は、基礎生物学研究所の大学院生養老瑛美子、寿崎拓哉助教、川口正代司教授らが中心となって、神戸大学の豊倉浩一研究員、深城英弘教授との共同研究として行われました。
[研究サポート]
本研究は、文部科学省科学研究費補助金、吉田育英会のサポートを受けて行われました。
[本件に関するお問い合わせ先]
基礎生物学研究所 共生システム研究部門
大学院生: 養老 瑛美子 (ヨウロウ エミコ)
Tel: 0564-55-7563 Fax: 0564-55-7563
E-mail: e-yoro@nibb.ac.jp
基礎生物学研究所 共生システム研究部門
教授: 川口 正代司 (カワグチ マサヨシ)
Tel: 0564-55-7564 Fax: 0564-55-7564
E-mail: masayosi@nibb.ac.jp

