基礎生物学研究所



2014.03.18
基礎生物学研究所 環境光生物学研究部門(皆川純教授、得津隆太郎助教)、スイス・ポールシェラー研究所(ガーガリー・ネギ研究員)、ハンガリー科学アカデミー(ギョーゾー・ガラブ科学アドバイザー)、フランス原子力代替エネルギー庁(ジョバンニ・フィナッチ研究部長)などの研究グループは、緑藻が光合成反応を調節するしくみ、ステート遷移の機構を明らかにしました。植物は光合成によって太陽からの光エネルギーを獲得しさまざまな生命活動を支えています。光合成は多くのステップからなる複雑な反応ですが、光エネルギーを捉えるステップには、光化学系I、光化学系IIと呼ばれる2つの色素タンパク質複合体(光化学系)が主要な役割を果たします。この2つの光化学系の連携は光合成反応全体の効率を左右する重要な問題です。この連携のコンセプトは“ステート遷移”呼ばれ40年以上前に発見されました。その後、多様な光合成生物がステート遷移を行っていることがわかり、その詳細をめぐっては多くの研究・議論が行われてきました。今回、これまでの常識を覆し、“アンテナ”タンパク質の移動が実際にはほとんど起きていないこと、そして“アンテナ”タンパク質の性質が変化していることを明らかにしました。これにより光エネルギーの効率的変換へ向け大きな足がかりが得られたことになります。この研究成果は、米国科学一般誌PNAS(米国科学アカデミー紀要)の電子速報版に3月17日以降(米国東部時間)に掲載されます。
【研究の背景】
植物や藻類によって光合成反応が行われる際、集めた光エネルギーは直列に並ぶ2つの光化学系(光化学系2/光化学系1)において化学エネルギーへと変換されます。この一連の反応を効率よく行うためには、2つの光化学系がバランス良く駆動される必要があります。このバランスが崩れると、活性酸素が発生するなど細胞に深刻なストレスがかかってしまうからです。自然の中では、季節や時間によって光の波長や強さが大きく変化するため、植物や藻類はこのアンバランスの危険に常にさらされています。しかし、このアンバランスはステート遷移と呼ばれるしくみによって常に補正されているため、通常は深刻な問題はおこりません。これまでの研究から提唱されてきたのは、「光化学系IIに光が集まりすぎると、光化学系IIの周辺に位置する光のアンテナタンパク質LHCIIがはずれてもう一方の光化学系(光化学系I)に結合し、今度はそこで光を集める」というモデルでした。このモデルはこれまで行われてきた試験管内実験の積み重ねから推測したもので、生きた細胞の中の姿が解析されたことはありませんでした。
【研究の成果】
研究グループは、ラウエ・ランジュヴァン研究所(仏グルノーブル)において加速器が作り出す中性子ビームを単細胞緑藻クラミドモナス細胞に当ててその小角散乱を測定し、ステート1では整然としていたチラコイド膜の規則構造がステート2では崩れること、細胞の円偏光吸収スペクトルを調べ、LHCIIの結合を含めた光化学系IIの全体構造がステート遷移の前後でほぼ保存されること、そして分光学的測定により光化学系IIの集光能力が大きく低下する一方光化学系Iの集光能力はわずかしか増強されないことを明らかにしました(図1)。また、クラミドモナスがステート遷移をおこすとき、光化学系IIに結合しているLHCIIの数は変わらないこと、一方の光化学系Iに結合する数は少数増えることを生化学的に明らかにしました(図1)。これらの結果、ステート遷移は従来考えられていたような「単純な光のアンテナの移動」ではなく、「光のアンテナの性質変化」であることがわかり、バランスの崩れた光化学系IとIIの連携は、それにより改善されていることが明らかになりました(図2)。
【成果の意義と今後の展開】
本研究により、従来型のモデルでは説明のつかなかった現象も説明され、水生光合成生物である藻類が周辺光環境に応じて2つの光化学系の連携を最適化するメカニズムの全体像が明らかになりました。驚くべきことに、そのメカニズムはこれまで陸上植物で提唱されてきた単純なものとは異なり、これまで全く別の現象とされてきたNPQ機構(過剰光エネルギー散逸機構)と酷似していることがわかりました(NPQ機構の詳細は2013年5月28日のプレスリリース「過剰な光エネルギーを消去する実体、光合成タンパク質超複合体を発見」を参照)。今後は2つのメカニズムの統一理解が進むものと思われます。代替エネルギー源として近年注目されている藻類ですが、大規模培養では日中の太陽光によるストレスへの対策が重要であることがわかってきました。本研究の成果はこうした応用面においても技術的な突破口を与えると期待されます。
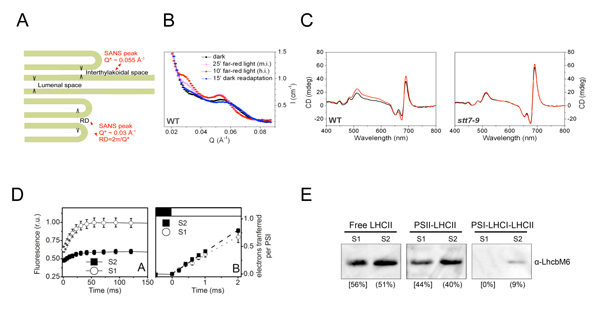
図1: 緑藻クラミドモナスのステート遷移解析
クラミドモナス細胞をステート1(S1、光化学系IIの集光能力>光化学系Iの集光能力)からステート2(S2、光化学系IIの集光能力<光化学系Iの集光能力)へ遷移させ、以下の解析を行った。 (A)中性子小角散乱実験で解析されるチラコイド膜構造の模式図、(B)中性子小角散乱によるブラッグピーク、(C)円偏光スペクトル測定によるpsi型ピーク(左:野生株、右:ステート遷移欠失変異株stt7)、(D)蛍光収率から見積もった光化学系II集光能力(左)とエレクトロクロミックシフト測定により見積もった光化学系I集光能力(右)、(E)各光化学系に物理的に結合しているLHCII量。
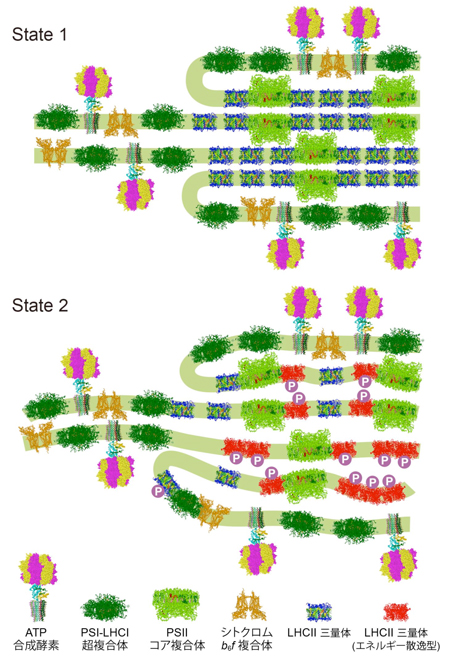
図2: 今回の研究から提唱されたステート遷移のモデル図
光化学系II(PSII)が大きな集光能力を持つ状態(ステート1)では、光のアンテナ(LHCII、青色)はPSIIコア複合体(黄緑色)の両側に結合し、葉緑体のチラコイド膜(緑色の帯)は整然と重なっています。青色光、強光、無酸素環境などによりPSIIにエネルギーが集まりすぎると、その状態を解消するためにLHCIIのリン酸化が起こり(丸で囲んだP)、葉緑体のチラコイド膜は波打ち重なりが乱れます。この時LHCIIは凝集体を作るなどしてエネルギー散逸型(赤色)となり、集めたエネルギーを捨て始めます。また、一部はPSIIから離れ光化学系I(PSI)に結合しPSIのアンテナとなります。結果として、PSIIの集光能力は小さくPSIの集光能力は大きく補正され、PSIIとPSIの連携状態は改善されることになります。
【発表雑誌】
◯ 雑誌名 : Proceedings of the National Academy of Sciences of the United States of America (米国科学アカデミー紀要)
◯ 論文タイトル : Chloroplast remodeling during state transitions in Chlamydomonas reinhardtii as revealed by non-invasive techniques in vivo。
(非侵襲技術により明らかになった、ステート遷移におけるクラミドモナス葉緑体のリモデリング)
◯ 著者 : Gergely Nagy 、 Renáta Ünnep 、 Ottó Zsiros 、 Ryutaro Tokutsu 、 Kenji Takizawa 、 Lionel Porcar 、 Lucas Moyet 、 Dimitris Petroutsos 、 Győző Garab 、 Giovanni Finazzi 、 and Jun Minagawa
【研究グループ】
本研究は、基礎生物学研究所環境光生物学研究部門(皆川純教授、得津隆太郎助教、滝澤謙二(元NIBBフェロー))、ハンガリー科学アカデミー、仏・原子力代替エネルギー庁、仏ラウエ・ランジュヴァン研究所による研究グループで実施されました。
【研究サポート】
本研究の一部は、内閣府最先端・次世代研究開発支援プログラム、科学技術振興機構戦略的創造研究推進事業、文部科学省植物CO2資源化研究拠点ネットワーク、NEDO戦略的次世代バイオマスエネルギー利用技術開発事業、科学研究費助成事業(研究活動スタート支援)のサポートを受けて行われました。
【本件に関するお問い合わせ先】
基礎生物学研究所 環境光生物学研究部門
教授: 皆川 純 (ミナガワ ジュン)
〒444-8585 愛知県岡崎市明大寺町字西郷中38
TEL: 0564-55-7515
E-mail: minagawa@nibb.ac.jp
ホームページ http://www.nibb.ac.jp/photo/
【報道担当】
基礎生物学研究所 広報室
TEL: 0564-55-7628
FAX: 0564-55-7597
E-mail: press@nibb.ac.jp

