自然科学研究機構 基礎生物学研究所
植物の光合成装置を過剰な光ストレスから守るNPQ反応を制御するタンパク質因子「DET1」が、ゲノム内の“動く遺伝子”であるDNAトランスポゾンのサイレンシングにも関与することが明らかになりました。DET1遺伝子が欠損した緑藻クラミドモナス変異株は、弱光条件下において当初はきわめて遅い成長を示しますが、やがてその成長は劇的に改善されます。この回復は、DNAトランスポゾン「
Bill」が光保護遺伝子
LHCSRの転写因子に自発的に挿入されNPQ反応を無効化することによって引き起こされたものでした。この発見は、緑藻細胞内において、LHCSRを介した短期的な環境応答能力と、トランスポゾン活性化による長期的なゲノム適応能力とが、DET1によって統合的に制御されているという、これまで知られていなかった新たな分子メカニズムの存在を示唆しています。本成果は、生物が環境の変化や変動に対してどのように適応し進化してきたのかという根源的な問いに、新たな視点を提供するものです。本研究は自然科学研究機構基礎生物学研究所環境光生物学研究部門の鎌田(藤村)このみ研究員と皆川純教授によって行われ、その成果は英国の専門誌
New Phytologist電子版(2025年8月7日付)に掲載されました。
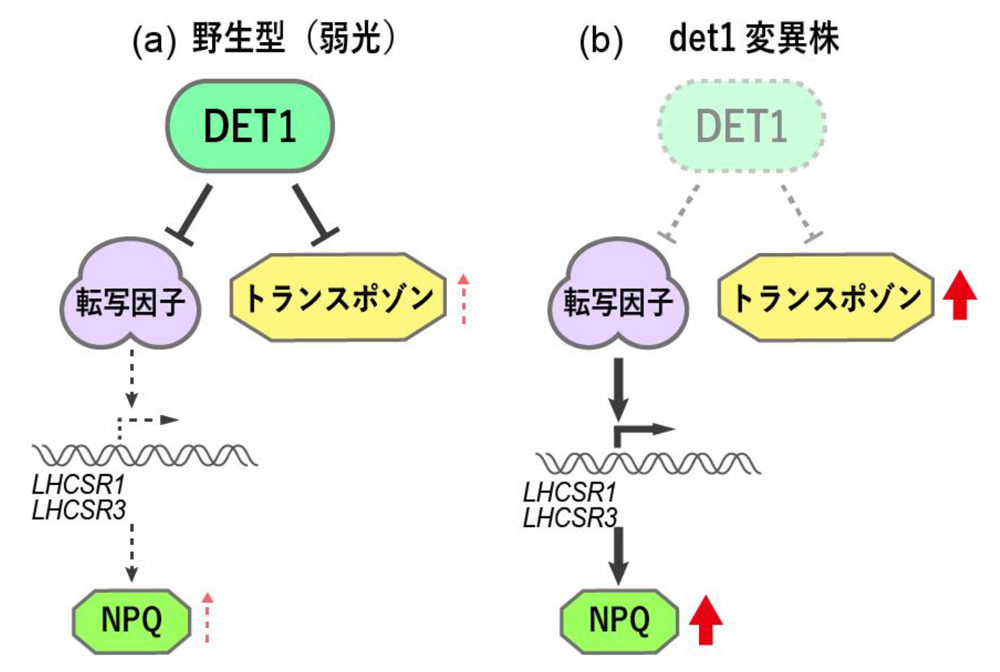
図:通常時(弱光)、野生株ではLHCSRの転写因子とトランスポゾンの活性はDET1によって抑制されていますが、DET1を欠損すると(
det1変異株)、LHCSRは転写されてNPQ反応が上昇するとともに、トランスポゾンの抑制が解除されます。
【研究の背景】
植物は、強すぎる太陽光にさらされると強光ストレスによって光合成装置が損傷する恐れがあります。これを防ぐため、植物は過剰な光エネルギーを熱として逃がすNPQ
*1(non-photochemical quenching)という防御機構を備えています。緑藻クラミドモナス
*2(
Chlamydomonas reinhardtii)では、NPQの活性化にはLHCSR
*3というタンパク質が関わっていますが、LHCSRが常に発現していると光エネルギーの利用効率が低下するという“副作用”があります。このためLHCSRの発現は、通常DET1
*4というユビキチンE3リガーゼ
*5によって抑制されています。DET1が欠損した変異体(
det1変異体)ではLHCSRが過剰に発現し、強い光の下では良く成長しますが、弱い光の下では成長が阻害されます(図1)。本研究は、
det1変異体を弱い光で培養した際に、その成長不良が自発的に回復する現象に注目しました。
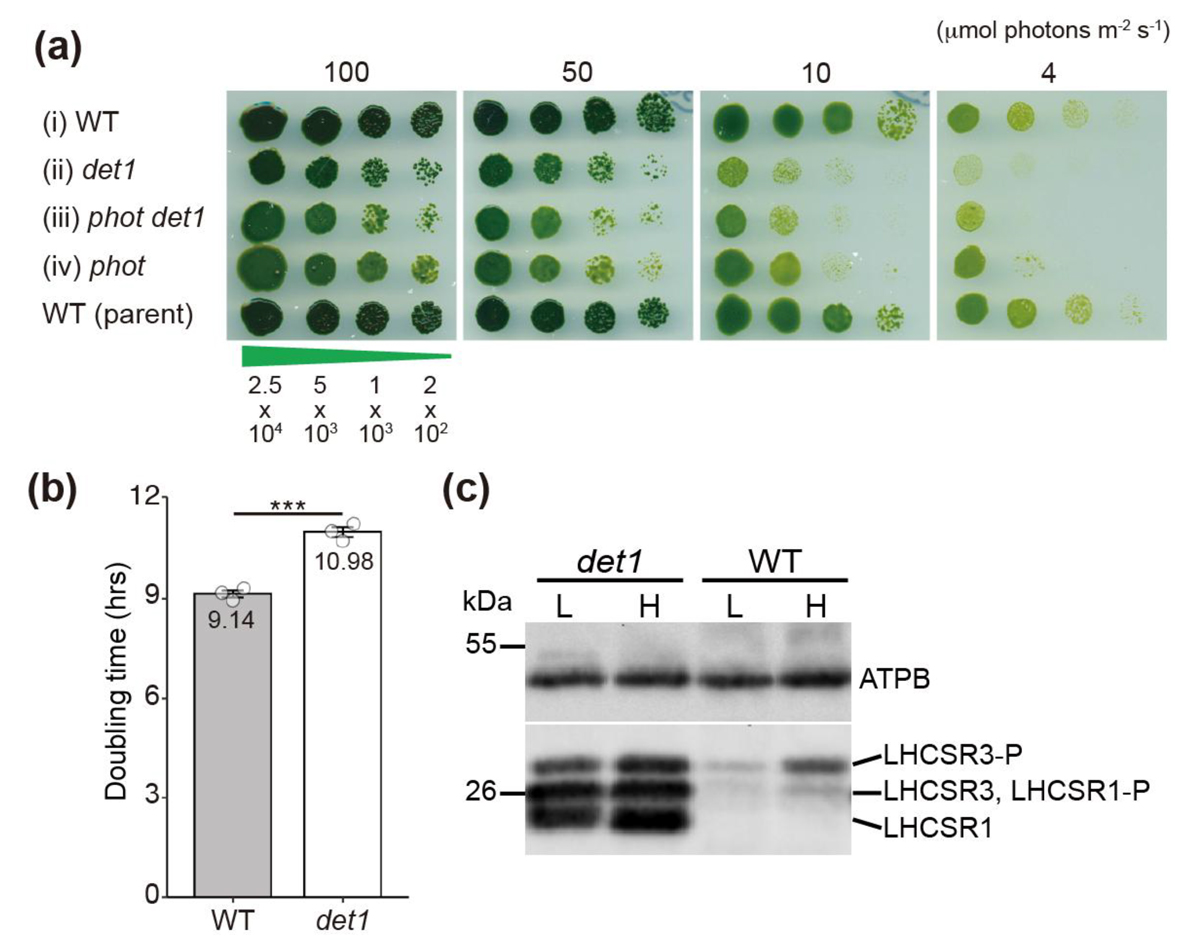
図1.
det1変異株の弱光条件下での成長阻害(a,b)とLHCSRの恒常的発現: 野生株(WT)と
det1変異株の成長比較(a)、倍加速度比較(b)およびLHCSRタンパク質(LHCSR1とLHCSR3)の発現レベル比較。DET1の欠損が弱光下での成長に与える影響と、その原因となるLHCSRの蓄積が示されている。L(弱光)、H(強光)。
【研究の成果】
本研究では、単細胞緑藻クラミドモナスを用いて、二つの重要な生物学的プロセス、すなわちNPQとトランスポゾン
*6活性化が、光ストレス下でどのように関連付けられているのかを明らかにすることを目指しました。
まず、
det1変異株を弱い光で培養した際に、同時多発的に現れる成長回復株「
DET-
ONE
SUPPRESSOR」(
dos変異体)を複数単離しました。詳細な遺伝学的解析と分子生物学的解析の結果、
dos変異株群では、
LHCSR遺伝子の転写因子である
CrCOおよび
NFYB*7に、「
Bill」と呼ばれる558塩基対のDNAトランスポゾンが挿入されていることがわかりました(図2) 。
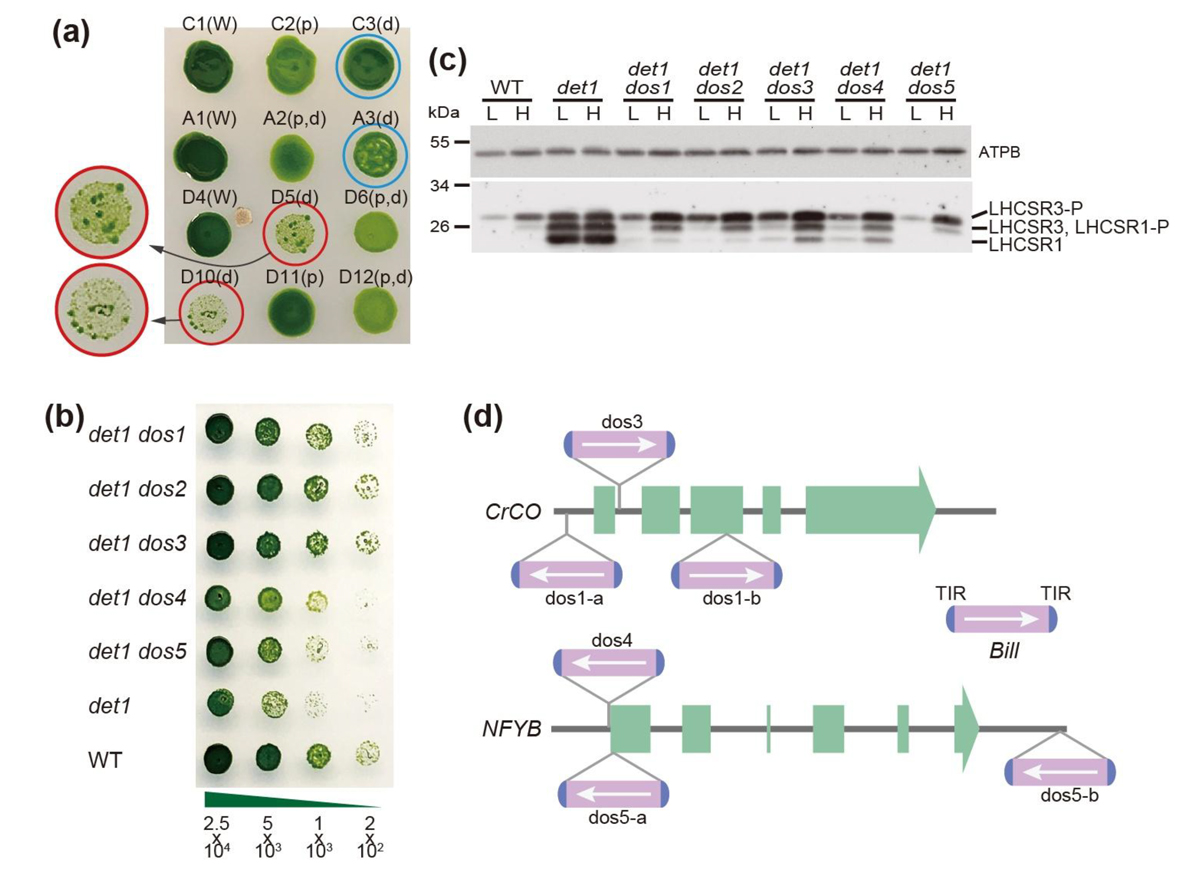
図2.(a)
det1変異株におけるトランスポゾン
Billの挿入と成長回復: 野生株(W)と
det1変異株第2世代(d)の弱光環境における成長。
det1変異株は基本的に野生株より遅い成長を示すが、青丸で囲った
det1変異株は野生株並の成長を示し、また赤丸で囲った
det1変異株の一部も野生株並の成長を示している。(b)単離した回復株
dos1〜
dos5の成長は
det1よりも明らかに速く、それは弱光条件でLHCSRの発現が野生株レベルに抑制されていたことによるものだった(c)。(d)それらのゲノムにはいずれも
LHCSRの転写因子である
CrCOや
NFYBにDNAトランスポゾン
Billが挿入されていた。
トランスポゾン
Billは、両端に特徴的な逆位反復配列(TIR)と標的部位重複配列(TSD)を持つが転移酵素自体は持たない、いわゆる「非自立型DNAトランスポゾン」です。
dos変異株では、
Billの挿入によって
LHCSR遺伝子の転写が抑制されるためNPQが誘導されずに効率の良い光合成反応が行われ、その結果弱光条件での生育が回復していたことがわかりました。トランスポゾンは、進化・適応には有用であるためストレス条件下などでは必要とされる一方でゲノムの不安定化要因でもあるため、通常時は沈静化されています。
det1変異株ではトランスポゾンが活性化された結果ゲノムに変異が加えられ(
dos変異)、その結果NPQの副作用が解決され弱光条件での生育が向上していたのです。驚くべきことに、植え継ぎ実験の結果は、
det1変異株では
Billの転移は現在も進行中であることを示していました(図3)。このことは逆に、平常時はDET1がトランスポゾンの転移を沈静化させてゲノムの安定性を維持している、言わば「ゲノムの守護者」として役割を果たしていることを意味しています。
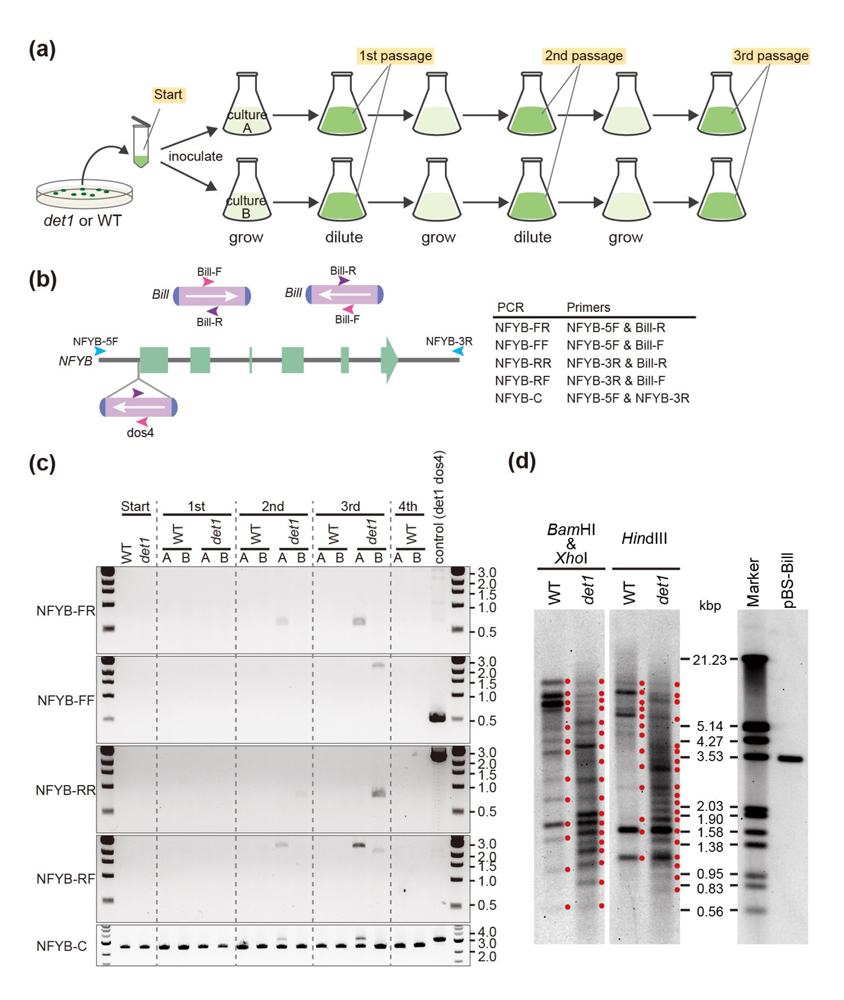
図3.
det1変異株における
Bill転移因子の活性化促進:
det1変異株と野生株におけるトランスポゾン
Billの転移頻度の比較。(a)野生株と
det1変異株を数世代植え継ぎ、そのゲノムに与える影響を調べた。(b)NFYB遺伝子に
Billが挿入された場合に検出できるPCR用プライマーの位置と(c)実際のPCR。A,Bは独立した2つの培養。(d)
Bill配列を用いたサザンブロッティングを行い、野生株と
det1変異株におけるゲノム上の
Bill配列の散らばりを比較した。赤点はバンドの位置を示す。
【本研究の意義と今後の展開】
本研究により、クラミドモナスにおいて、DET1が光ストレス応答とゲノム内のDNAトランスポゾン(
Bill)の活性化を制御するという二重の役割を担っていることが明らかになりました(図4)。DET1は陸上植物やヒトにも存在します。シロイヌナズナでは光形態形成に関わる因子として知られていますが、エピジェネティックな制御に関わるとの報告もあります。クラミドモナスと陸上植物は離れた種ですが、DET1を介したストレス応答とエピジェネティック制御機構は共通祖先から受け継がれた普遍的な戦略である可能性が示唆されます。一方で、ゲノム構造はDET1によって直接制御されているのではなく、DET1の下流因子によって間接的に制御されている可能性も考えられます。例えば、ヒトのアルツハイマー病由来細胞では、DET1が制御しているがん原性転写因子c-Junのレベルが上昇するとトランスポゾンが活性化するとの報告があります。クラミドモナスにはc-Junはありませんが、c-JunのようなDET1制御下の転写因子がトランスポゾン活性化に関与しているのかもしれません。
今回の「光ストレス応答とトランスポゾン活性化の統合制御」という発見は、生物の環境適応メカニズムに新たな視点を提供します。強光ストレスという環境からの「挑戦」に対して、それが「天気の変化」のような短期的なものであればNPQ反応のような一過的な応答で対応し、同時に「気候変動」のような長期のものである可能性を想定しトランスポゾンの活性も開放しておく、という生物種としての生存戦略があるのかもしれません。今後、強光ストレスだけではなく他のストレス条件でも同様の仕組みが存在するのか、他の生物種にも同様の仕組みが存在するのかといった研究の進展が期待されます。また、この研究は、光防御とゲノム変異誘発の両面から植物の耐環境性能を高める技術開発にも貢献すると考えられます。
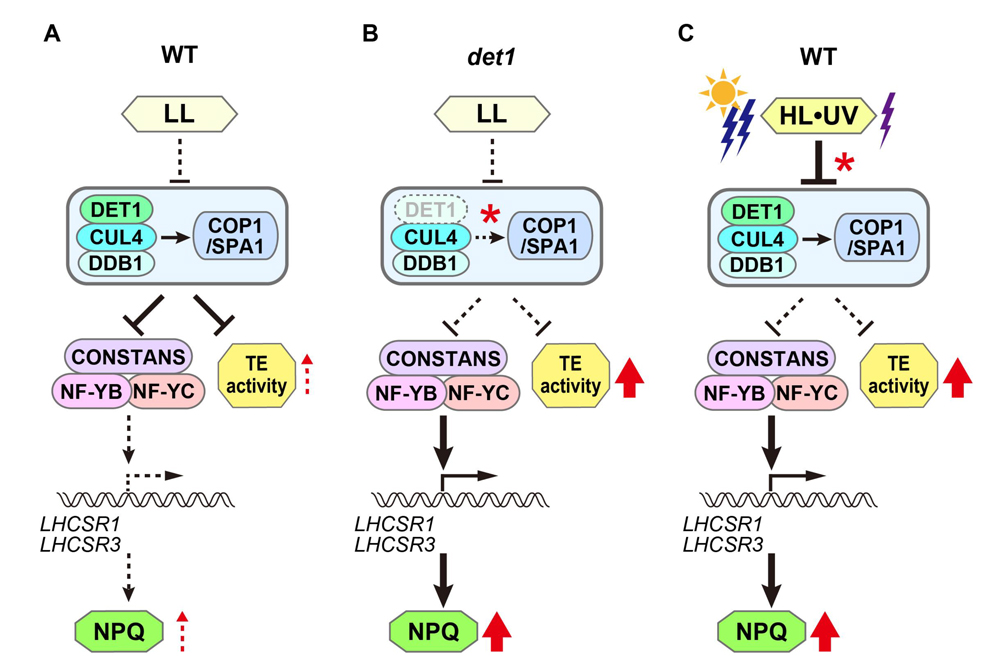
図4. DET1による光ストレス応答と転移因子活性化の統合制御モデル: DET1が光防御(NPQ)とゲノムの可塑性(トランスポゾン活性化)を統合的に制御しているかを野生株(弱光)、
det1株(弱光)、野生株(強光)に分けて示した。DET1の他に制御に関与する可能性のあるユビキチンE3リガーゼ因子(COP1,SPA1,CUL4,DDB1)、LHCSRの転写因子(CONSTSNS,NFYB,NFYC)を示した。赤*はその後の制御が分岐するチェックポイントを示す。
【用語解説】
1. NPQ(Non-Photochemical Quenching, 光化学系非光化学的消光): 植物が過剰な光エネルギーを熱として散逸させ、光合成装置を損傷から守る防御機構です 。
2. クラミドモナス(
Chlamydomonas reinhardtii): 本研究で用いられた単細胞の緑藻で、光合成研究のモデル生物として広く利用されています 。
3. LHCSR(LIGHT-HARVESTING COMPLEX STRESS RELATED protein): 光ストレス応答時に誘導される光防御タンパク質で、NPQの活性化に中心的な役割を果たします 。
4. DET1(DE-ETIOLATED1): 植物の光形態形成や光防御に関わるタンパク質で、ユビキチンE3リガーゼ複合体の一部として機能します。本研究では、光防御と転移因子の活性化抑制という二重の役割が明らかになりました 。
5. ユビキチンE3リガーゼ: タンパク質にユビキチンという小さな分子を結合させ、そのタンパク質の分解を促進する酵素複合体です。細胞内の様々なタンパク質の量を調節しています 。
6.トランスポゾン(Transposon)もしくは転移因子(Transposable Elements, TEs): ゲノム内を移動できるDNA配列で、「動く遺伝子」とも呼ばれます。ゲノムに多様性をもたらし、進化に貢献すると考えられています 。トランスポゾンには、RNAを介するRNAトランスポゾンと介さないDNAトランスポゾンがあり、
BillはDNAトランスポゾンです。
7. CrCO/NF-Ys(CONSTANS / NUCLEAR FACTOR-Y B and C subunits): LHCSRの発現を促進する転写因子複合体です 。
【掲載誌情報】
雑誌名:
New Phytologist
掲載日: 2025年8月7日
論文タイトル: From Photoprotection to Plasticity: Transposon Activation in the
Chlamydomonas det1 Mutant
著者: Konomi Fujimura-Kamada and Jun Minagawa
DOI:
https://doi.org/10.1111/nph.70436
【研究サポート】
本研究は、科学研究費補助金(学術変革領域研究「光合成ユビキティ」(23H04960)、基盤研究(S) (21H05040))のサポートを受けて行われました 。
【問い合わせ先】
基礎生物学研究所 環境光生物学研究部門
教授 皆川 純(ミナガワ ジュン)
〒444-8585 愛知県岡崎市明大寺町字西郷中38
TEL: 0564-55-7515
E-mail: minagawa@nibb.ac.jp
ホームページ:
https://www.nibb.ac.jp/photo/
【報道担当】
基礎生物学研究所 広報室
TEL: 0564-55-7628
FAX: 0564-55-7597
E-mail: press@nibb.ac.jp




