自然科学研究機構 基礎生物学研究所
情報・システム研究機構 国立遺伝学研究所
カブトムシのオスは「角」でメスを巡り争い、オスのスズムシは翅に備わる「発音器官」を奏してメスを誘います。このように、昆虫類の形態には、繁殖の成功に基づく淘汰の結果として、様々な雌雄差(性的二型)を認めます。昆虫類の性的二型は、
doublesexと呼ばれる遺伝子で制御されることが知られています。完全変態類昆虫の
doublesexは、雌雄で異なるスプライシング調節を受け、ひとつの遺伝子にもかかわらず、性的二型形成に関する遺伝子の転写を雌雄で正反対に調節して、雌雄それぞれの特徴の形成を制御します。昆虫類以外の節足動物の研究から、
doublesexは、進化の過程で、元々オスの形態形成を促進していましたが、後にメスの形態形成を促進する機能を獲得したと推定されています。
doublesexが如何にメスの形態形成への機能を獲得したかという疑問は、動物の性に関する謎の一つです。
今回、基礎生物学研究所 進化発生研究部門の千頭康彦研究員(元・総合研究大学院大学 大学院生)と新美輝幸教授、久留米大学の奥野未来助教、国立遺伝学研究所の豊田敦特任教授、東京工業大学の伊藤武彦教授からなる研究グループは、昆虫進化の初期に出現したシミ類に着目して
doublesexの機能を解析し、その祖先状態を推定しました。その結果、シミ類マダラシミの
doublesexは雌雄で異なるスプライシング調節を受けますが、メスの形態形成への機能をもたないことが明らかになりました。一方、マダラシミの
doublesexは幾つかの遺伝子の発現をメスで促進することが分かりました。これらの結果は、
doublesexは昆虫進化の初期段階で既に雌雄で異なるスプライシング調節を受け、メスに特異な幾つかの遺伝子の発現を促進する機能をもち、そして完全変態類出現後にメスの形態形成に対する機能を獲得した可能性が高いことを示唆しています。では、
doublesexのどのような変化がメスの形態形成への機能と関連したのでしょうか。本研究では、メスの形態形成に対する機能をもつ種のDoublesexタンパク質に特有なアミノ酸配列を発見しました。この結果から、本研究は、アミノ酸配列の変更によって
doublesexは新機能を獲得したとする仮説を提唱しました。本研究成果は、有翅昆虫類と昆虫類以外の節足動物との間にあった知見のギャップを埋めることに成功し、
doublesexの特殊な進化史を新たに推定するものです。
本成果は、日本時間2022年7月13日付で「
Molecular Biology and Evolution」誌に掲載されました。
【研究の背景】
動物の多くはオスとメスが交配し、次世代たる子を得ます。オスとメスという性は、ほとんどの動物門で知られており、動物の進化の歴史で一度獲得されたことが示唆されています。単一起源の可能性が高いにもかかわらず、動物の性を決定し雌雄差をもたらす分子メカニズムは、分類群間や近縁種間、同種内の集団間でさえも異なる場合があります。動物の性決定メカニズムにみられる顕著な多様性は、進化生物学の関心の対象として、その進化過程が精力的に研究されてきました。
動物の性は様々な遺伝子による数珠つなぎの連鎖反応(性決定カスケード)を通じて決定されます。これまでの研究で注目されてきた性決定メカニズムの多様性は、性決定カスケードを構成する性決定遺伝子の種間での違いでした。一方で、昆虫類の研究では、同一の性決定遺伝子における機能の多様性も明らかになっています。昆虫類の性決定遺伝子
doublesex(
dsx)は雌雄で異なる選択的スプライシング(用語1)を受けて、最終的にオスとメスで異なるアミノ酸配列をもつタンパク質が合成されます(図1)。ハチ類を除く完全変態類(用語2)では、雌雄それぞれに特異的なDsxタンパク質は性的二型の形成に関わる標的遺伝子の転写を雌雄で正反対に調節することで、雌雄それぞれの特徴の形成を促進します。すなわち、
dsxはひとつの遺伝子ながらも雌雄で正反対の機能を担うという特色をもちます。最近、ハチ類や幾つかの不完全変態類の
dsxも同様に雌雄で異なる配列のタンパク質を指定しますが、興味深いことに、オスの特徴の形成のみにかかわり、メスの特徴の形成には必要ないことが報告されていました。
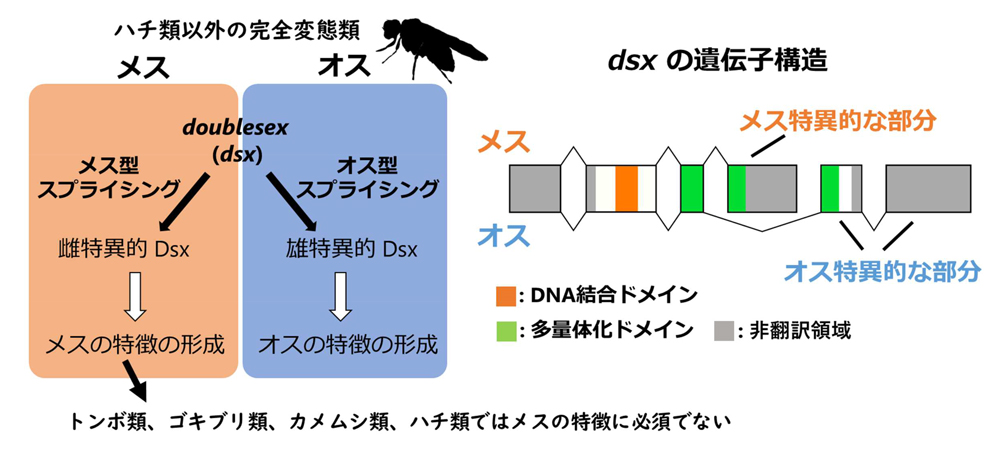 図1.昆虫類における性決定遺伝子doublesexの制御様式と遺伝子構造。
図1.昆虫類における性決定遺伝子doublesexの制御様式と遺伝子構造。雌雄で異なるスプライシング制御によって雌雄で異なる配列のタンパク質が最終的に生じます。
現在、昆虫類の
dsxは雌雄で異なるスプライシングによる制御とメスの特徴の形成への機能を段階的に獲得したという仮説が提示されています。しかしながら、これまでの不完全変態類の知見は少数の種からの断片的な情報であり、また昆虫類と昆虫類以外の節足動物との間にも知見のギャップがありました。そのため、
dsxがメスの発生に対する機能をどのようにして獲得したのか、という問いは大部分が不明のままとなっていました。
【研究の成果】
本研究では、これまでの知見のギャップを解消し、
dsxの機能の進化過程を推定するため、昆虫類の
dsxの系統関係を調査し、また昆虫進化の初期段階に分岐したシミ類(用語3)の
dsxの機能を解析して昆虫類における
dsxの祖先状態を推定しました。さらに、公共データベースなどを駆使して、昆虫類における
dsxの配列を大規模に比較しました。
まず、昆虫類における
dsxの分子進化の全体像を把握するために、公共データベースなどを利用して昆虫類32目のうち29目から
dsxの配列を入手し、分子系統解析を行ないました。その結果、昆虫類の
dsxは昆虫進化の初期に重複していたことが明らかになりました。すなわち、昆虫類の共通祖先は2つの
dsxをもっていたことが示唆されました。さらに、今回の解析では、2つの
dsxのうちひとつはほとんどの昆虫類で存在していましたが、もう一方はシミ類、カゲロウ類、ナナフシ類のみで認められ、他のグループでは失われている可能性が示されました。この一部の昆虫類に認められる
dsxを
dsx-likeと名付けました。この結果や昆虫類の系統関係を踏まえると、シミ類は
dsxと
dsx-likeを両方もつという昆虫類の祖先状態を維持し、かつ昆虫類と昆虫類以外の節足動物との知見のギャップを埋めるのに適した題材であることが示唆されました。
そこで、本研究ではシミ類のマダラシミ
Thermobia domestica(図2A)に着目して
dsxの解析を行ないました。
dsxの配列決定並びに幼虫期のRNA干渉(RNAi)法による機能解析の結果、本種の
dsxは雌雄で異なるスプライシング調節を受けていること(図2B)、オスの形態形成には寄与するがメスの形態形成には必須でないこと(図2C)が分かりました。尚、
dsx-likeは
dsxと異なり、性特異的スプライシング調節を受けず、雌雄の形態形成のいずれにも必要ありませんでした。
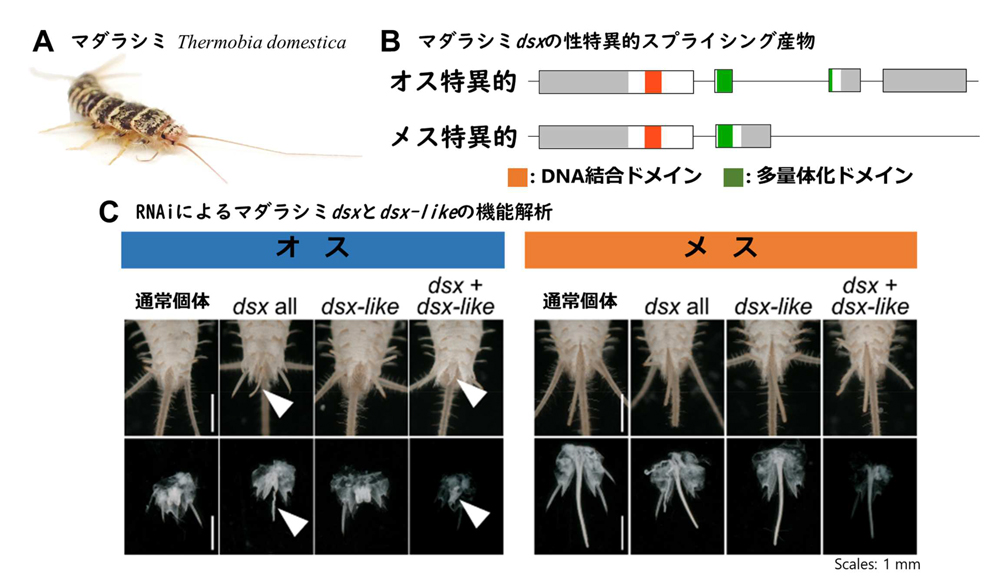 図2.マダラシミにおけるdoublesexのスプライシング産物の構造と機能。
図2.マダラシミにおけるdoublesexのスプライシング産物の構造と機能。機能解析では、幼虫期を通じて
dsx或いは
dsx-likeの機能を阻害し、オスの外部生殖器とメスの産卵管を観察しました。矢尻はオスにもかかわらず、
dsxの機能阻害によって生じた産卵管状の構造を表わしています。メスの産卵管は、
dsxや
dsx-likeの機能が阻害されても正常に形成されました。図中の
dsx all、
dsx-like、
dsx +
dsx-likeはそれぞれ
dsx、
dsx-like、
dsxと
dsx-like両方の機能阻害個体を示しています。
続いて、形態以外への機能も解析したところ、予想外なことに、本種の
dsxと
dsx-likeの機能の阻害により、通常メスだけで発現している卵黄タンパク質前駆体
vitellogenin 遺伝子の発現量がオスで上昇し、メスで減少すること(図3)が明らかになりました。これらの結果は、本種の
dsxが形態形成に対してはオスのみで機能する一方で、形態形成以外の側面ではメスでも機能をもつことを示しています。この結果と甲殻類や有翅昆虫類からの研究の比較から、
dsxは4億年以上前の昆虫進化の初期段階で性特異的な選択的スプライシング調節を受け、形態形成以外でのメスに特異的な幾つかの遺伝子の発現を制御する機能を獲得し、3億年ほど前のハチ類以外の完全変態類の出現以降にメスの形態形成に対する機能を新たに獲得したという段階的な進化を辿った可能性が示唆されました。
では、
dsxのどのような変化がメスの形態形成への機能と関連したのでしょうか。昆虫類49種のDsxアミノ酸配列を比較し祖先配列を推定した結果、Dsxタンパク質がハチ類以外の完全変態類の共通祖先においてカルボキシル末端側(用語4)に16アミノ酸からなるモチーフを獲得していたことを発見しました(図4)。この結果から、本研究では、カルボキシル末端側へのモチーフの付加が
dsxにメスの形態形成に対する機能の獲得に関連したとする仮説を新たに提案しました。
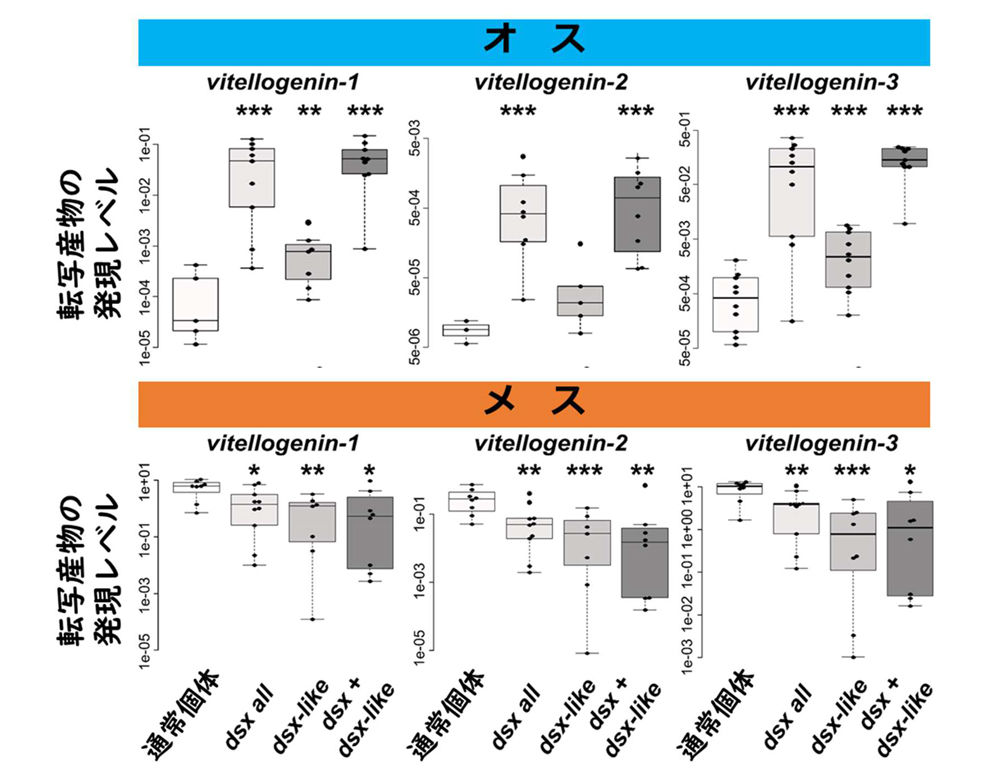 図3.マダラシミdoublesexのvitellogenin発現への機能。dsx
図3.マダラシミdoublesexのvitellogenin発現への機能。dsxや
dsx-like の機能を阻害した個体での
vitellogeninの発現を解析しました。マダラシミは少なくとも3つの
vitellogenin遺伝子をもちます。アスタリスクは通常個体との間に統計学的に有意な差が検出されたRNAi処理区を表わしています。
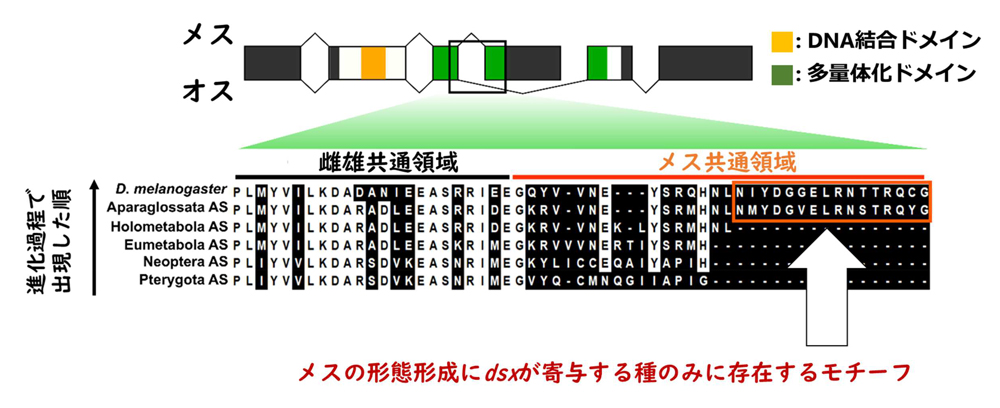 図4.doublesexの配列比較と祖先配列推定。
図4.doublesexの配列比較と祖先配列推定。
【今後の展望】
今回の成果により、昆虫類の形態の雌雄差を担う
dsxの進化仮説をアップデートすることに成功しました(図5)。この成果は、選択的スプライシング調節を受ける遺伝子が如何にして新たな機能を獲得するのか、という真核生物で幅広くみられる現象ながらこれまで研究の進んでこなかった問いに貢献すると期待されます。
今後、メスの形態形成に
dsxが必須でない種におけるDsxのカルボキシル末端側に、今回発見したモチーフを実験的に導入することで、今回提唱した仮説を検討していくことになります。
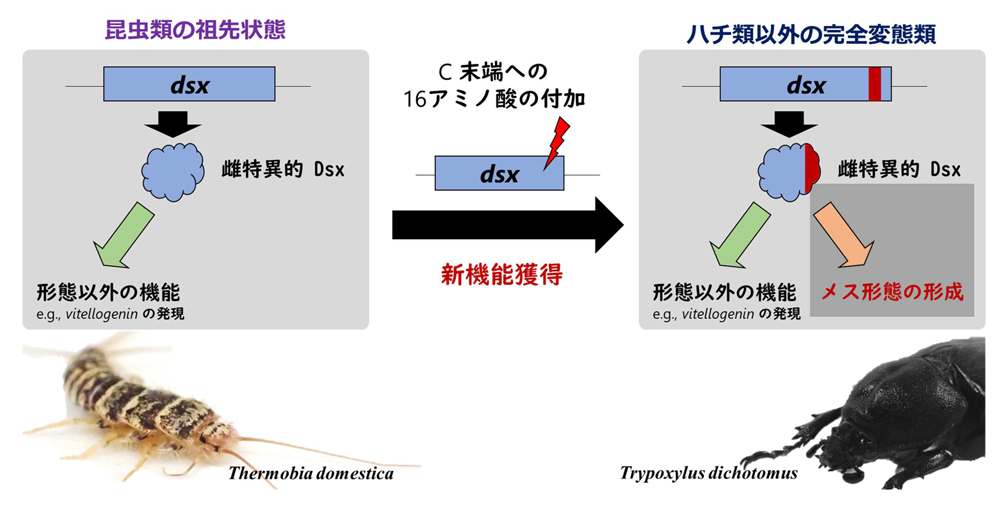 図5.本研究で推定されたdoublesexの機能進化過程。
図5.本研究で推定されたdoublesexの機能進化過程。昆虫類の祖先状態を保持していると推定される代表としてマダラシミ
Thermobia domestica、ハチ類以外の完全変態類の代表例としてカブトムシ
Trypoxylus dichotomusを示してある。
【用語】
1.選択的スプライシング:ゲノムDNA上の遺伝子は、伝令RNAに情報が写し取られ、さらに伝令RNAの情報を基にタンパク質が合成されることで機能します。真核生物の遺伝子は、伝令RNAに写しとられる部分(エクソン)と、写しとられない部分(イントロン)が組み合わさった構造をしています。伝令RNAがつくられる際には、エクソンとイントロンの情報をもった未成熟な前駆体がまず作られます。その後、イントロンの部分が取り除かれて成熟した伝令RNAが作られます。このイントロンを取り除きエクソン同士をつなぎ合わせる過程はスプライシングと呼ばれます。スプライシングの過程で、異なるエクソンが組み合わさり配列の異なる伝令RNAが生じることがあります。このような異なるエクソンを組み合わせ、単一の遺伝子から異なる配列をもつ伝令RNAを生じるスプライシングは特別に「選択的スプライシング」と呼ばれます。選択的スプライシングは、ひとつの遺伝子から複数種の伝令RNAを生じ、最終的に機能の異なるタンパク質への翻訳につながります。このことから、選択的スプライシングは、限られた遺伝子数の中で真核生物が極めて複雑かつ多様な特徴をもつことができる一因とされます。
2.完全変態類:幼虫から蛹を経て成虫へと至る成長過程は完全変態と呼ばれます。完全変態は昆虫類の進化の道筋の中で一度だけ生じたとされています。完全変態という特徴を共有する昆虫のグループは完全変態類と呼ばれます。この10年ほどの研究で、完全変態類は3憶5000万年以上前に出現したとされます。最初に分岐した完全変態類はハチの仲間であることが示唆されています。ハチ類を除く完全変態類の仲間はAparaglossataとして纏められます。
3.シミ類:シミ類は成虫でも翅をもたない無翅昆虫であり、かつ翅を獲得した有翅昆虫類の姉妹群にあたるグループです。このグループは、生涯を通じて脱皮し続け、幼虫と成虫の形態の変化もほとんどない無変態という過程で成長します。その系統学的な位置や無翅・無変態などといった昆虫類の共通祖先がもっていたとされる数多くの特徴を今なおも維持していることから、昆虫進化研究の鍵となるグループです。性に関しても、ダーウィンがシミ類には性差がないと言及しているように、本種の性差は卵巣や精巣を除けば、メスの産卵管とオスの外部生殖器にほぼ限局され、さらに交尾をしないという昆虫類の祖先的な特徴を維持しています。日本では外来種も含めると17種が知られ、本研究で用いたマダラシミは本邦の南西諸島や欧米諸国の家屋内に生息しています。
4.カルボキシル末端:タンパク質は数多くのアミノ酸が連なったポリペプチド鎖から構成されます。ポリペプチド鎖にはアミノ末端(N末)とカルボキシル末端(C末)という2つの末端が存在します。アミノ酸はアミノ基(-NH
2)とカルボキシル基(-COOH)をもつ化合物であり、アミノ基とカルボキシル基が数珠つなぎに結合することでポリペプチド鎖を構成します。アミノ末端は他のアミノ酸と結合していない遊離したアミノ基のある側で、カルボキシル末端は遊離したカルボキシル基のある側です。カルボキシル末端には、しばしばタンパク質の機能に重要なドメインがあります。Dsxもカルボキシル末端に他のタンパク質との結合に必要なドメインをもっています。
【発表雑誌】
雑誌名:Molecular Biology and Evolution
掲載日:July 12th, 2022, 21:00 (UTC)
論文タイトル: Evolutionary history of sexual differentiation mechanism in insects
著者:Yasuhiko Chikami, Miki Okuno, Atsushi Toyoda, Takehiko Itoh, Teruyuki Niimi
DOI:
https://doi.org/10.1093/molbev/msac145
【研究グループ】
本研究は、基礎生物学研究所を中心に、東京工業大学、国立遺伝学研究所、久留米大学が参画した共同研究チームによって遂行されました。
【研究サポート】
本研究は、科学研究費助成事業(JP25660265, JP16H02596, JP16H06279 [先進ゲノム解析研究推進プラットフォーム(PAGS)])と日本科学協会の笹川科学研究助成による支援を受けて遂行されました。
【本研究に関するお問い合わせ先】
基礎生物学研究所 進化発生研究部門
教授 新美輝幸
TEL: 0564-55-7606
E-mail: niimi@nibb.ac.jp
【報道担当】
基礎生物学研究所 広報室
TEL: 0564-55-7628
FAX: 0564-55-7597
E-mail: press@nibb.ac.jp




