基礎生物学研究所



2016.09.15
植物は,光のエネルギーを利用して二酸化炭素を固定し糖を合成します(光合成)。そのため、強い光の方がより光合成をすると考えられがちですが、実際には強すぎる光は光合成装置を壊してしまいます。この危険を避けるため、植物は強い光を浴びたときには、そのエネルギーを熱に変換してわざと逃がすガス抜きのしくみを発達させました。qEクエンチング(*1)と呼ばれる、このブレーキ役のしくみは、環境が変動する中で植物が生き残るために必要であったと考えられています。これまでqEクエンチングの詳細は謎に包まれていましたが、今回、これまで光合成とは直接関係ないと思われていた青色光受容体の一つフォトトロピン(*2)が決定的な役割を果たしていることが明らかになりました。その結果、これまで個別の現象と考えられていた、青色光の受容、光合成、光防御が実は分子レベルで繋がっていることになり、環境変化がおきた際の細胞中の一連の反応の流れの全体像が見えてきました。
今後は、生育環境が整った圃場では、より光合成を進める側に反応のバランスを傾ける、あるいは砂漠地帯の池のような過酷な環境でエネルギー藻類を培養する場合は、より光合成を抑える側に反応のバランスを傾けるなど、光合成反応調節技術への発展が期待されます。
本研究は基礎生物学研究所の皆川純教授、得津隆太郎助教と、フランス国立科学研究センターのジョバンニ・フィナッチ博士らを中心とした国際共同研究チームによる成果です。
本研究成果は,英国の科学誌『Nature』の電子先行版(2016年9月14日付)に掲載されました。
【研究の背景】
植物細胞の中には、葉緑体という細胞内小器官(オルガネラ)があります。葉緑体の中にはチラコイドと呼ばれる膜でできた袋構造があり、その表面にはたくさんの光合成装置が並んでいます。ここに光があたると、光合成反応のアクセルが踏まれ、電子が流れATPができます。この電子とATPを使って二酸化炭素を固定し糖分を作る、その一連の反応が『光合成』です。光合成はいわば光からエネルギーを取り出す反応ですから、より多く光を当てた方がより光合成をするものと考えられがちですが、実際には強すぎる光を当てると光合成装置は壊れてしまいます。この危険を避けるため、植物は強い光を浴びたときには、そのエネルギーを熱に変換してわざと逃がすガス抜きのしくみを発達させました。いわば光合成にブレーキをかけるしくみを備えたのです。一見無駄にみえるこの光合成のブレーキ役はqEクエンチングと呼ばれます。qEクエンチングは水中の植物である藻類からコケ植物、さらには陸上植物にいたるまでほとんどの植物に備わっていることがわかっており、環境が変動する中で植物が生き残るために必要な機能であったと考えられています。
これまで、光合成反応の中でも、いかに光が集められ、いかに電子が流れ、いかにATPが作られるのか、といった光合成のアクセルに関してはたくさん研究が行われてきました。しかし、qEクエンチングの詳細は長い間謎に包まれてきました
【研究の成果】
研究グループは、単細胞植物とも言うべき緑藻クラミドモナスにおいて、葉緑体の中の光合成装置に結合してqEクエンチングを引き起こすLHCSR3タンパク質に注目しました(LHCSR3タンパク質の詳細は2013年5月28日のプレスリリース「過剰な光エネルギーを消去する実体,光合成タンパク質超複合体を発見」を参照)。細胞に強い光を当てた時にLHCSR3が合成されることは知られていましたが、“光が強い”ことを細胞がどのようにして知るのかこれまでわかっていませんでした。まず、基礎生物学研究所の地階に設置されている「大型スペクトログラフ(図1)」を使い、何色の光の刺激によってLHCSR3が合成されるかを調べました。その結果、LHCSR3の合成、そしてそれによって起こるqEクエンチングには、青色光や赤色光を吸収するクロロフィルが必要なこと、そしてさらに青色光を受容する別の因子も必要であることがわかったのです(図2)。qEクエンチングは、起こりすぎた光合成にブレーキをかけるというフィードバック機構ですから、クロロフィルが必要であることは予想できたものの、青色光受容体が必要であったことは驚きの発見でした。これまで、植物の持つさまざまな現象において(光屈性や葉緑体運動、花芽形成、気孔開閉、概日リズムなど)、クリプトクロムやフォトトロピンといった青色光受容体が関与することが知られていましたが、それらは光合成反応に直接かかわるものではないとされていたからです。研究グループは、さらにこれらの青色光受容体が失われた変異株を利用して研究を続けました。その結果、フォトトロピンを失った変異株ではLHCSR3が合成されないことを突き止めました(図2)。つまり、光合成装置を強い光から守るqEクエンチングには、過剰な光合成活性と青色光受容体であるフォトトロピンの両方が必要であることがわかったのです。
【本研究の意義と今後の展開】
青色光は可視光の中で最もエネルギーが高いので、吸収分子により最も大きな影響を与えることができます。また、水中では赤色光はすぐに減衰しますが青色光は比較的保たれるため、他の色よりも深くまで届くという点でも有利です。本研究で示された青色光を利用する光合成のブレーキ機能は、水圏で生活する藻類やコケ植物が、急に光が強くなったことを感知する際に特に有効だったのだろうと研究グループは考えています。強光を浴びた陸上の植物もqEクエンチングをおこしますが、そのqEクエンチングをおこすタンパク質はフォトトロピンの影響を受けません。その代わりに、フォトトロピンは葉緑体そのものが光から逃げる反応(葉緑体運動)に関わっています。葉緑体運動は、qEクエンチングを介して光合成反応自体を抑制するわけではありませんが、葉緑体が光を避けた結果として細胞全体の光合成活動にブレーキがかかります。そのため、水圏でも陸上でも、フォトトロピンは光合成にブレーキをかけていることになります。太古の昔、植物は水中で生活していました(藻類)、それが進化して陸上に上がったのが現在の陸上植物であるとされています。今後は、今回明らかになった光合成のブレーキ機能が水中から陸上へいかに進化したのか、またなぜそれが必要であったのか等の理解も深まっていくことでしょう。
本研究により、これまでそれぞれ別の現象であると考えられてきた、フォトトロピンによる青色光の受容、クロロフィルによる光合成、そしてqEクエンチングによる光防御、この三者が分子レベルで繋がっていることがわかり、環境変化がおきた際に細胞中で起こる反応の流れの全体像が明らかになりました(図3)。直射日光はヒトにとって辛いものですが、植物にとっても辛いため、直射日光が一年中降り注ぐような環境は植物にとって大きな負担になっています。今後は、生育環境が整った耕作地で作物を育てる場合は、より光合成を進める側に反応のバランスを傾ける、あるいは砂漠地帯の池のような過酷な環境でエネルギー藻類を培養する場合は、より光合成にブレーキをかける側に反応のバランスを傾けるなど、光合成反応調節技術への発展が期待されます。
*1 qEクエンチング:クエンチング(q)とは一般に(火などが)“消える”、(乾きを)“癒やす”などの意味で使われるが、光合成反応においては、光エネルギーを吸収して興奮状態になったクロロフィルが鎮まることを意味する。その際の分子機構の違いにより、qEクエンチング、qTクエンチング、qMクエンチングなどが知られている。qEクエンチングは、光合成装置に電子が流れ葉緑体にエネルギー(E)が与えられたときに生じるフィードバック型のクエンチングのことで、その研究は今世紀の光合成研究におけるトレンドの一つになっている。
*2 フォトトロピン:植物に広く存在する「青色光受容体」タンパク質の一つ。青色光受容に関わるフラビン色素団を持つLOV(light-Oxgygen-Voltage)ドメインとその信号を次の信号伝達タンパク質へ伝える役割を担うキナーゼドメインからなる。陸上植物では、光屈性、葉緑体運動、気孔開閉などに関与することが知られている。

図1.基礎生物学研究所の地階に設置された「大型スペクトログラフ」。30 kWの光源が250~1000
nmに分光され、全長約10メートルの馬蹄型の焦点曲面に投射される。この施設を利用して、クラミドモナス細胞を各ポジションに置き、それぞれに異なる強い単色光を照射してLHCSR3の合成やqEクエンチングの強度を調べた。
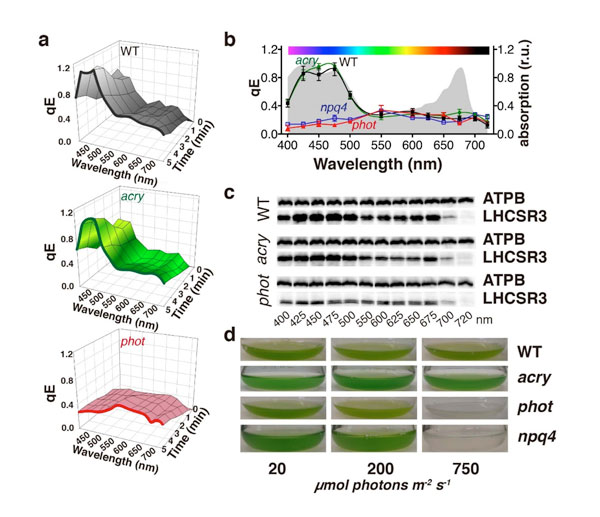
図2.大型スペクトログラフによって得られたアクションスペクトルはLHCSR3合成とqEクエンチングにフォトトロピンが関与していることを示した。a. 各波長を指定時間照射した際のqEクエンチング量。WT, 野生株; acry, クリプトクロム欠失変異株; phot, フォトトロピン欠失変異株。b. 各波長を4時間照射した際のqEクエンチング量。c. 各波長を4時間照射した際のLHCSR3量。d. 野生株と変異株を弱光、中光、強光(20, 200, 750 μmol photons m-2 s-1)条件で培養した際のクラミドモナス細胞の生育の様子。npq4, LHCSR3欠失変異株。フォトトロピンを欠失した株(phot)やLHCSR3を欠失した株(npq4)は強光環境では生育できないことがわかる。
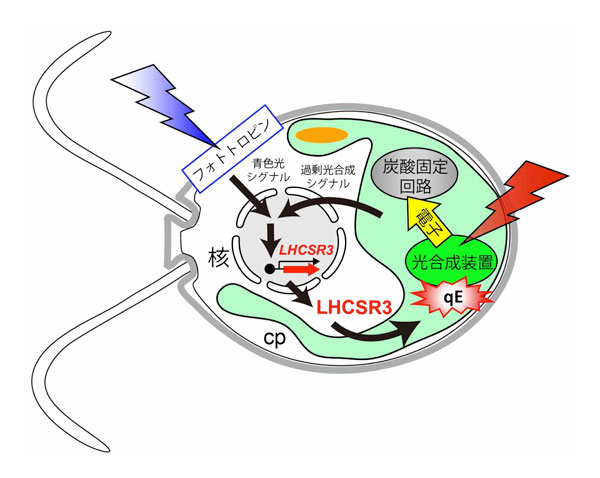
図3.本研究によって明らかになった光合成の負のフィードバックのモデル図。1)強い青色または強い赤色の光を葉緑体の中の光合成装置が吸収すると、過剰な光合成が起こり、そのシグナルが細胞質に伝わる。2)弱い青色光を青色光受容体フォトトロピンが吸収すると、そのシグナルが細胞質に伝わる。この2つのシグナルがLHCSR3遺伝子の発現には必要である。合成されたLHCSR3が葉緑体に輸送され光合成装置に結合すると、光合成装置は光を捨てはじめ(qEクエンチング)、光合成はそれ以上進行しない。
【掲載誌情報】
雑誌名: Nature(英国9月14日オンライン公開、9月22日号掲載)
論文タイトル: A blue-light photoreceptor mediates the feedback regulation of photosynthesis
(青色光受容体が光合成のフィードバックを引き起こす)
著者: Dimitris Petroutsos*, Ryutaro Tokutsu*, Shinichiro Maruyama, Serena Flori, Andre Greiner, Leonardo Magneschi, Loic Cusant, Tilman Kottke, Maria Mittag, Peter Hegemann, Giovanni Finazzi & Jun Minagawa
http://dx.doi.org/10.1038/nature19358
【研究グループ】
基礎生物学研究所の皆川純教授、得津隆太郎助教と、フランス国立科学研究センターのジョバンニ・フィナッチ博士らを中心とした国際共同研究チームによる成果です。
【研究サポート】
本研究は、基礎生物学研究所、文部科学省、科学技術振興機構、新エネルギー・産業技術総合開発機構(NEDO)のサポートを受けて行われました。
【本研究に関するお問い合わせ先】
基礎生物学研究所 環境光生物学研究部門
教授 皆川 純(ミナガワ ジュン)
〒444-8585 愛知県岡崎市明大寺町字西郷中38
TEL: 0564-55-7515
E-mail: minagawa@nibb.ac.jp
ホームページ http://www.nibb.ac.jp/photo/
助教 得津 隆太郎(トクツ リュウタロウ)
TEL: 0564-55-7517
E-mail: tokutsu@nibb.ac.jp
【報道担当】
基礎生物学研究所 広報室
TEL: 0564-55-7628
FAX: 0564-55-7597
E-mail: press@nibb.ac.jp

