植物の害虫として身近な存在であるアブラムシ(アリマキ)は、自身の体内に「ブフネラ(
Buchnera aphidicola)」と呼ばれる細胞内共生細菌を保持しています。アブラムシは、餌である植物の篩管液に不足しているアミノ酸などの栄養供給を、この細菌に完全に依存しています。ブフネラは「バクテリオサイト」という専用の細胞に格納され、母から子へと厳密に受け継がれています。アブラムシとブフネラの関係は約2億年に及ぶ極めて安定したものですが、一部の系統では、ブフネラに加えて別の細菌を必須とする「多重共生系」の例が知られています。このような系では、ブフネラは一貫して保持されている一方で、もう片方のパートナーである細菌が、新しく参入した細菌に役割を明け渡し、最終的に入れ替わってしまうことがあります。しかし、新しい細菌がどのようにして既存の共生システムに入り込み、地位を確立したのか、そしてそれが共生システム全体にどのような影響を与えたのか、具体的なプロセスは謎に包まれていました。
今回、基礎生物学研究所 進化ゲノミクス研究室の野崎友成助教、小林裕樹研究員(研究当時 現東京農業大学)、重信秀治教授を中心とするグループは、日本に生息するクリオオアブラムシ(
Lachnus tropicalis)が持つ多重共生系を詳細に解析しました。その結果、新しく参入した共生細菌であるセラチア(
Serratia symbiotica)A系統が、以下の3つのプロセスを経て共生系を再構築していることを発見しました。
1. 「居抜き物件」としての継承:先代細菌(セラチアB系統)が利用していた細胞ニッチや伝達ルートをそのまま継承していた。
2. 代謝システムの統合:ブフネラとアブラムシとの相補的な代謝システムに、ゲノムレベルで統合されつつあった。
3. 共生細菌ゲノムの最適化を駆動:パートナーの置換とそれに続く再構築が、居残ったブフネラのゲノムのさらなる「スリム化」をも促進させていた。
また、詳細なゲノム解析の結果、新参のセラチアA系統は、自由生活を送る近縁な祖先型セラチアと同程度の数の遺伝子をもち、多くの必須アミノ酸やビタミンの合成能力を保持していることがわかりました。これらの遺伝子が、ブフネラが持っていない代謝経路をカバーしていた一方で、ブフネラが得意とするアミノ酸代謝経路では偽遺伝子が多く検出され、両者が代謝レベルで機能を補完し合っていることがわかりました。
さらに、セラチアB系統を保持する、ヨーロッパ産の近縁種(
Lachnus roboris)のブフネラも含めて比較ゲノム解析を行った結果、クリオオアブラムシの共生系では、セラチアB系統からA系統への置換イベントの後に、ブフネラは特定の代謝経路(トリプトファン合成経路など)を失い、その機能が新参セラチア(セラチアA系統)によって補完されていることが判明しました。この結果は、パートナーの交代がシステム全体の再編成を引き起こし、ブフネラのゲノムのさらなる機能の特殊化(縮小進化)を駆動した可能性を示しています。
本研究は、アブラムシの多重共生系におけるパートナーの置換が、既存の関係性の崩壊と再構築において重要な進化的要因であることを強く示しています。本成果により、長期間維持されてきた共生システムが、新たなパートナーの参入をきっかけとしてダイナミックに更新され、共進化のあり方を劇的に変化させうることが示されました。
本研究成果は、2026年2月18日付で国際学術誌「
Proceedings of the Royal Society B: Biological Sciences」誌に発表されました。
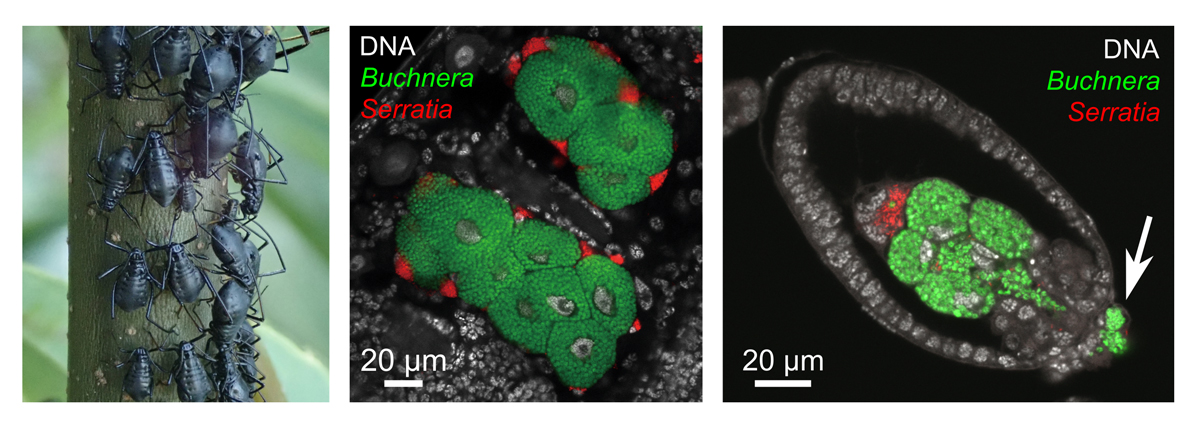
左図:クリオオアブラムシ、中央・右図:FISH法によって可視化された共生細菌(ブフネラとセラチア)のアブラムシ体内での局在(中央)と胚発生時の垂直伝達(右)(矢印は胚への侵入開始点を示す)。
研究グループはまず、蛍光
in situ hybridization (FISH)法による顕微鏡観察により、ブフネラとセラチアA系統からなる多重共生系を持つクリオオアブラムシの共生システムについて詳細な解析を行いました。そして、クリオオアブラムシに近縁な種の共生系(ブフネラとセラチアB系統からなる多重共生系)に関する過去の文献を紐解き、組織学的特徴を詳細に比較しました。その結果、この新参セラチア(セラチアA系統)は、先代の細菌(セラチアB系統)専用であったシース細胞(バクテリオサイトに近接する扁平な細胞)にそっくりそのまま局在していました。母親から子へ細菌を送り込むための伝達経路、そしてアブラムシの発生過程での共生器官への分配様式も、先代が利用していたものと全く同じであるとわかりました。これは、新参者が宿主共生系の「既存インフラ」をそのまま利用することで、速やかに必須のパートナーとしての地位を確立したことを示唆しています。
【研究の背景】
生物の進化において、異なる種が互いに利益を得ながら生きること(双利共生)は、新たな機能獲得を促す決定的な役割を果たしています。アブラムシの仲間は、植物の篩管液をエサとしていますが、これは糖が豊富である一方で、アブラムシにとって必須なアミノ酸やビタミンをほとんど含んでいません。アブラムシは不足する栄養の供給を、体内に保持する共生細菌「ブフネラ(
Buchnera aphidicola)」に完全に依存しています。一方でブフネラは、長い共進化の過程でアブラムシとの生活に徹底的に適応(特殊化)した結果、多くの自由生活に必須な遺伝子を失い、ゲノムサイズも0.6 Mbと非常に小さくなっています(一般的な細菌のゲノムサイズは4-5 Mb)。ブフネラはアブラムシの外では生きてゆけず、アブラムシはブフネラなしでは生きていけません。両者は、お互いがお互いの生存に必須な「運命共同体」であると言えます。この関係性の起源は古く、約2億年前に始まったと考えられています。
多くのアブラムシでは、ブフネラのみが必須な存在として保持されていますが、一部の系統では、ブフネラに加えて別の細菌を取り込み、生存に不可欠なパートナーとして定着させた「多重共生系」を確立しています(図1)[注釈1]。これまでの研究から、こうした多重共生系では、新しく参入した細菌が以前からいた細菌と置き替わったり、機能を補完し合ったりすることが知られていました。
しかし、「新参の細菌がどのようにして、不可欠な役割を担うに至るのか」「新たに再構築された共生系では何がどのように変化するのか」という具体的なプロセスについては、大きな謎として残されていました。というのも、共生細菌の置換イベントは進化の歴史の中では、痕跡が検出されることは多いものの、比較的稀なイベントであるからです。近縁なアブラムシ種の間で、細菌が入れ替わる前後を比較できるような「ちょうどよい」モデルはほとんど見つかっていませんでした。
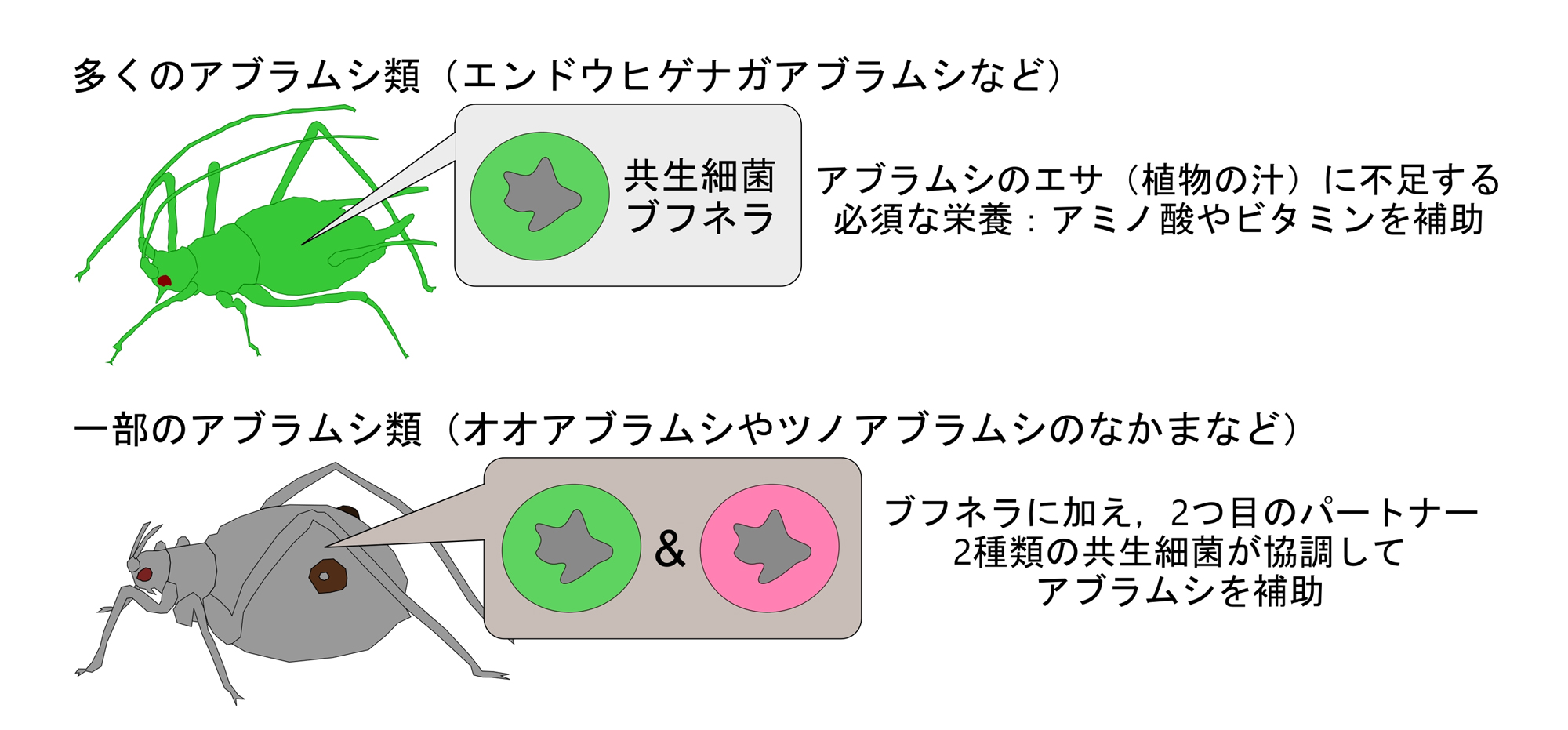 図1.アブラムシにおける必須共生細菌との関係性
図1.アブラムシにおける必須共生細菌との関係性
シンプルなブフネラだけの共生系と、2つの共生細菌が存在する多重共生系の例。
【研究の成果】
今回、研究グループは、メタゲノム解析と組織学的な観察を駆使し、日本に生息するクリオオアブラムシ(
Lachnus tropicalis)(図2左)の共生システムについて詳細な解析を行いました。本種は、ブフネラに加えて「セラチア(
Serratia symbiotica)」を体内に共生させていることが過去の研究から既に知られていました [注釈2]。また、ヨーロッパに生息する近縁種(
Lachnus roboris)を含むグループではセラチアの「系統B」が、クリオオアブラムシを含むアジアに生息するグループの派生的な種ではセラチアの「系統A」が保持されていることも分かっていました。これらのことから、クリオオアブラムシの共生系は、ブフネラと、比較的新参者である系統Aのセラチアで構成されていることが分かります(図2右)。研究チームは、既知の知見とクリオオアブラムシの事例を比較することで、セラチアのB系統からA系統への置換イベントに伴う、共生システムの組織学的・ゲノム学的な変化を捉えることに成功しました。
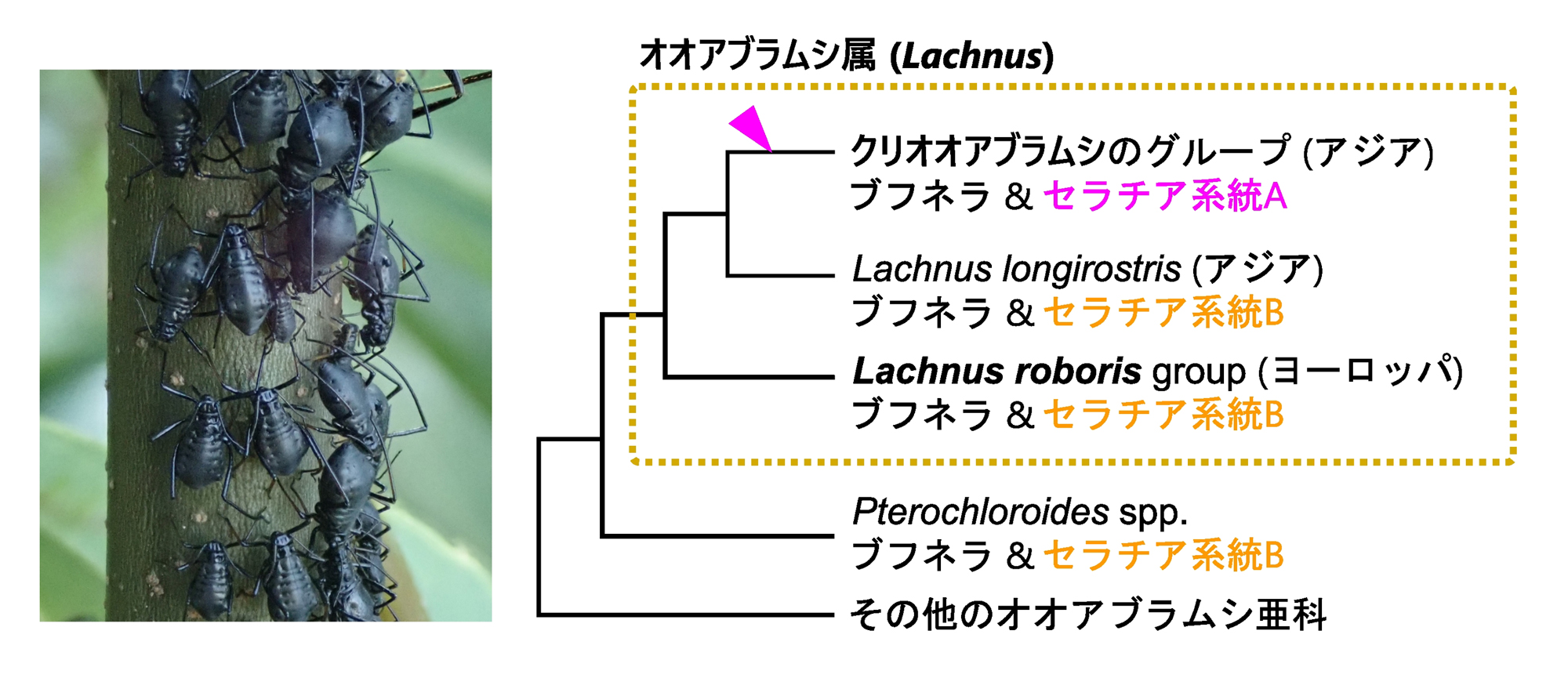 図2.アラカシ枝上のクリオオアブラムシ(左)と属内における共生細菌の遷移仮説(右)
図2.アラカシ枝上のクリオオアブラムシ(左)と属内における共生細菌の遷移仮説(右)
オオアブラムシ属の中で、系統樹の端に来る派生的なグループのみが、セラチアA系統を保持している。マゼンタの矢じりはセラチアA系統が獲得されたと考えられるタイミングを示す。
1. 共生インフラの「居抜き」利用を組織学的観察から発見
まず、セラチアA系統を保持するクリオオアブラムシの共生細菌伝達様式について、細菌の16S rRNAを標的とした蛍光
in situ hybridization (FISH)法による顕微鏡観察により詳細に調べました。そして、セラチアB系統を保持する
Lachnus roborisの共生細菌伝達様式を報告した文献を紐解き、両者を詳細に比較しました。その結果、クリオオアブラムシのセラチアA系統(以下、セラチアLt)は、セラチアB系統と同じく「シース細胞」という特定の共生細胞内に存在していることが判明しました(図3)。さらに、研究チームの得た観察結果は、母親から次世代へ細菌を送り込むための伝達経路や、胚発生の過程における共生器官への分配様式までもが、B系統をもつアブラムシでの観察と驚くほど一致していました(図4)。これは、新参の細菌が、宿主側が先代と確立していた既存の「共生インフラ(細胞ニッチや伝達ルート)」をあたかも「居抜き物件」のようにそのまま使いこなすことで、速やかに必須のパートナーとしての地位を確立したことを強く示唆しています。
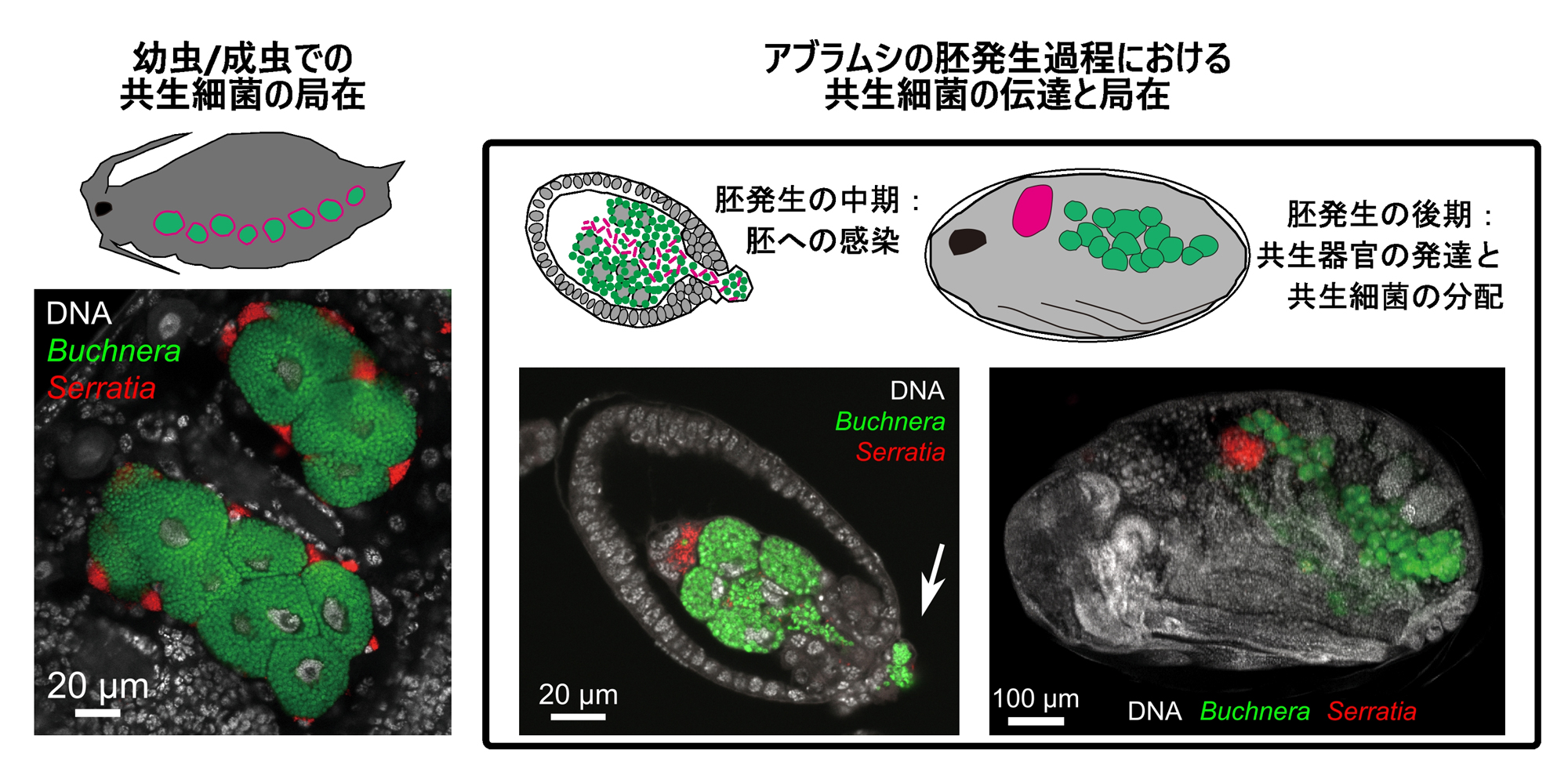 図3.クリオオアブラムシにおける共生細菌(ブフネラとセラチアLt)の細胞ニッチと伝達経路
図3.クリオオアブラムシにおける共生細菌(ブフネラとセラチアLt)の細胞ニッチと伝達経路
左:アブラムシの共生器官を構成する2種類の細胞のうち、ブフネラは巨大な「バクテリオサイト」を、セラチアLtはその周りに存在する扁平な「シースセル」を占有。右:アブラムシ胚発生過程において、ブフネラとセラチアLtは同じタイミングで胚へと感染(矢印は侵入開始点を示す)。その後の胚発生段階において、共生細菌たちは新たに形成された共生器官細胞へと分配される。FISH法により可視化。
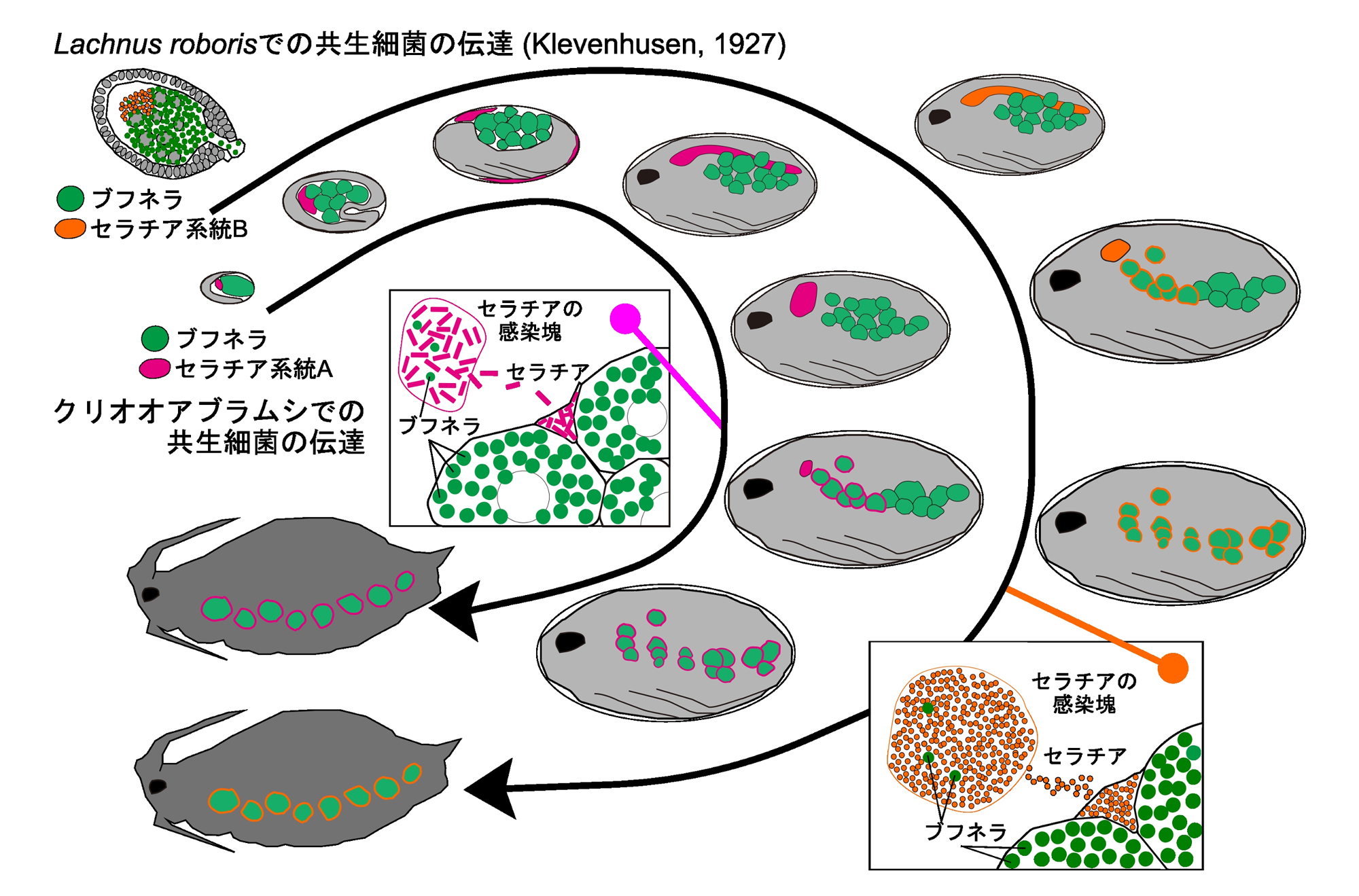 図4.クリオオアブラムシとその近縁種の胚発生過程における共生細菌伝達様式
図4.クリオオアブラムシとその近縁種の胚発生過程における共生細菌伝達様式
本研究におけるクリオオアブラムシでの観察(ブフネラとセラチアLt)と、ヨーロッパの近縁種である
Lachnus roborisにおける記載(Klevenhausen 1927)を比較した結果、セラチアの系統が異なるにかかわらず、アブラムシ体内での局在と垂直伝達様式には差異が認められなかった。
2. ゲノム解析から新参セラチアの機能と「分業」の確立を解明
次に、ゲノム解析[注釈3]によってセラチアLtの機能を調べたところ、そのゲノムサイズは約2.8 Mbであり、これは自由生活を送る近縁な祖先型(他種アブラムシの腸内に生息し、ゲノムサイズは約3.5 Mb)と比べてやや小さい値でした。またセラチアLtのゲノム上には、多くの必須アミノ酸やビタミンの合成にかかわる遺伝子が保持されていた一方で、アブラムシの細胞内での生活では重要性が低い、細胞壁合成やべん毛形成などにかかわる遺伝子では突然変異が蓄積しており、機能不全(偽遺伝子化)に陥っていました。これは、他の昆虫でもしばしばみられる「新参者の共生細菌」の特徴とよく一致しています。特筆すべきことに、セラチアLtでは、ブフネラやホスト側が主として担う代謝経路(ロイシンやメチオニン)の多くを放棄していることも分かりました。これらの結果は、セラチアLtがこの共生システムに参入したあと、新たにブフネラおよびアブラムシとの密接な機能補完を確立した、もしくは、しつつあることを示しています(図5)。
3. 比較ゲノム解析により見えてきた置換イベントとブフネラゲノムの縮小進化
さらに、セラチアの置換イベントが共生パートナーであるブフネラに与えた影響を評価するため、ヨーロッパの近縁種(
Lachnus roboris,セラチアB系統を保持)のブフネラゲノムとの比較解析を実施しました。その結果、クリオオアブラムシのブフネラでは、必須アミノ酸であるトリプトファンの合成経路が消失していることが明らかになりました。なお、この経路はセラチアLtによって完全にカバーされていました(図5)。これらの結果は、B系統からA系統へとセラチアの置換が起きた後に、セラチアだけでなく、ブフネラ側の機能も再編され、重要でなくなった経路に対する選択圧が緩和されたことで、ゲノムの縮小進化がさらに推し進められたことを示しています。
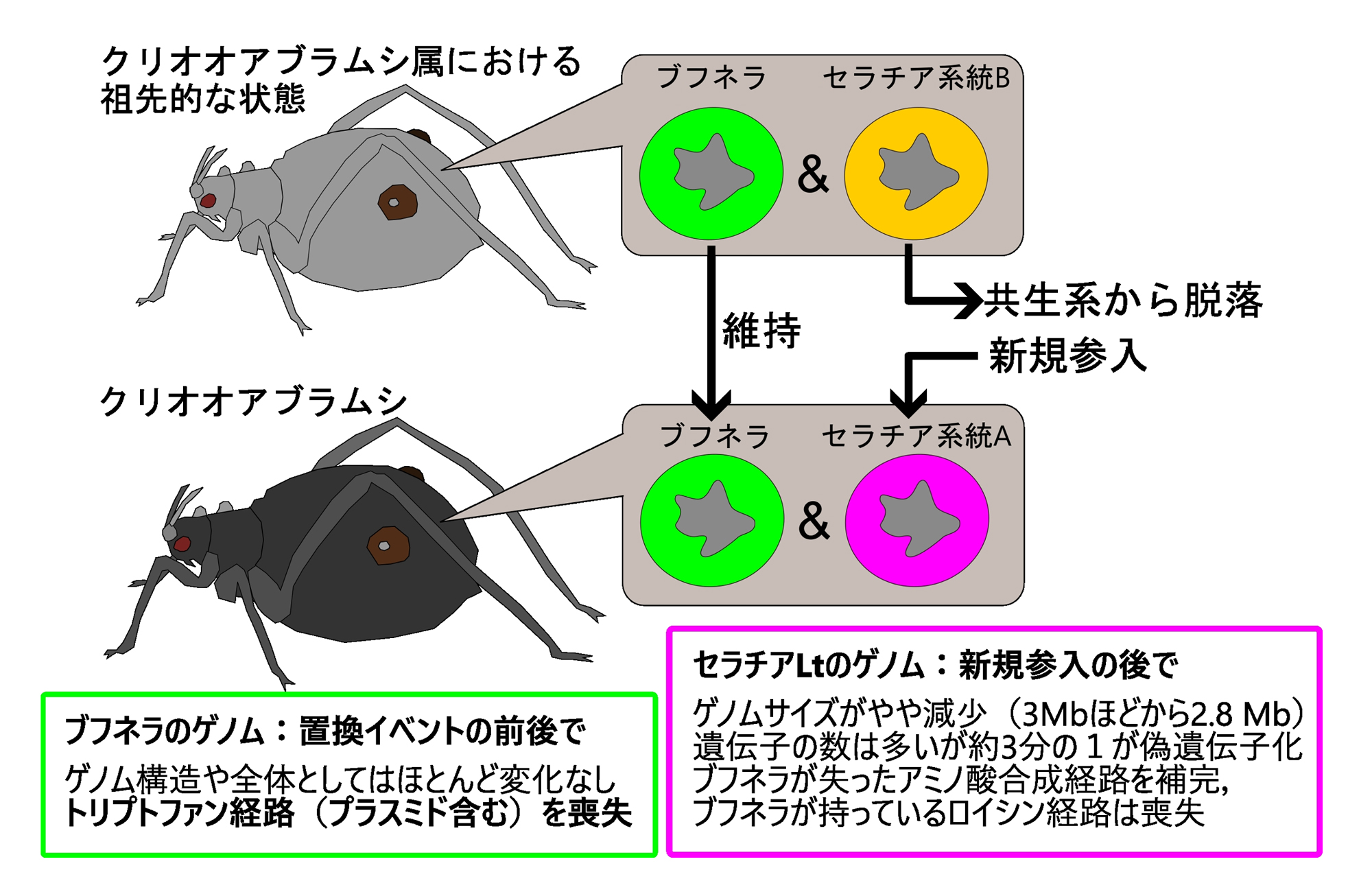 図5.オオアブラムシ属における共生細菌セラチアの置換と共生細菌ゲノムの変化
図5.オオアブラムシ属における共生細菌セラチアの置換と共生細菌ゲノムの変化
セラチアB系統の置換を契機とした共生システムの再構築の過程で、セラチアLt(系統A)だけでなく、ブフネラもさらなるゲノムの特殊化(縮小進化)を経験したと考えられる.
【今後の展望】
本研究では、クリオオアブラムシにおいて、新参共生細菌による「共生ニッチの居抜き」がシステム全体の機能を再編成し、一貫して存在しているブフネラ側の共進化をも促進していることを明らかにしました。これは、長期間にわたり極めて安定的に見えるアブラムシと細菌の共生関係において、その内実ではパートナーの交代劇や、それに伴う共生システムの再編成といったダイナミックな変化が絶えず起こっている可能性を示しています。
今後、特に注目すべきなのは、新参のセラチアが既存のシステムへどう「割り込んだか」という点であると考えています。共生細菌とホストの遺伝子発現解析(トランスクリプトーム解析)を通じ、セラチアLtの遺伝子のうち、ブフネラやホストの機能と重複する部分の発現がどのように制御・抑制されているのかを解明する予定です。この「機能のオーバーラップと抑制」という遺伝子レベルのダイナミズムは、既存の共生系を機能不全にせず、新たなパートナーを受け入れ、最終的にシステムを更新していく「置換イベント」の鍵を握っていると考えています。
一方で、「なぜこのような置換イベントが起きたのか」という生態学的な謎も残ります。一つの可能性として、ホスト昆虫の食性の変化や分布域の拡大といった背景が、新参のセラチアA系統を、従来のB系統よりも効率的なパートナーとして成立させたのかもしれません[注釈4]。興味深いことに、クリオオアブラムシを含む、A系統のセラチアを保持しているグループはアジアにおいて急速な種の多様化(適応放散)を遂げたことが分かっています。セラチアA系統の獲得および共生システムの再編性が、アブラムシの生存戦略にどう寄与し、適応放散のトリガーとなったのか、これは今後検証すべき、魅力的な仮説です。
今後は、より多くの近縁種の共生システムの解析を進めるとともに、分布や寄主植物などの、それぞれのアブラムシ種の生活史データと統合することで、置換イベントがアブラムシの適応進化に果たした真の役割を解明する予定です。本研究は、共生パートナーの交代劇を通じて新たな関係性が現れるというプロセスを明らかにしました。これは、共生系のダイナミックな本質に迫る重要な知見です。
【発表論文情報】
雑誌名:
Proceedings of the Royal Society B
掲載日: 2026年2月18日
論文タイトル: Symbiont replacement and subsequent genome erosion reshape a dual obligate aphid symbiosis
著者:Tomonari Nozaki, Yuuki Kobayashi, Mika Ikeda, Shuji Shigenobu
DOI:
https://doi.org/10.1098/rspb.2025.2484
【研究グループ】
本研究は、基礎生物学研究所の野崎友成助教、小林裕樹研究員(研究当時)、池田弥華技術支援員(研究当時)、重信秀治教授により実施されました。
【研究サポート】
本研究は、以下の文部科学省科学研究費補助事業(科研費)の支援を受けて行われました。
-
特別研究員奨励費「昆虫の菌細胞内共生系におけるホスト細胞・共生者の倍数化制御機構とその生物学的意義」(19J01756)
-
若手研究「昆虫における組織特異的な倍数化のメカニズムと生物学的意義の解明」(22K14901)
-
若手研究「昆虫共生細菌における倍数化の意義と制御機構の解明」(25K18554)
-
基盤研究B「共生系の遺伝子ネットワークの制御機構とその進化」(17H03717)
-
新学術領域研究(研究領域提案型)「進化の制約と方向性 ~微生物から多細胞生物までを貫く表現型進化原理の解明~」(17H06384)
-
基盤研究A「アブラムシ細胞内共生の分子機構をゲノム編集で明らかにする」(20H00478)
【用語解説および補足】
[注釈1] 進化ゲノミクス研究室では、以前からアブラムシの多重共生系に着目して研究をすすめている。2022年にはササコナフキツノアブラムシ
Ceratovacuna japonicaの実験室飼育系を確立し、ブフネラとアルセノフォナス(
Arsenophonus)による多重共生系を詳細に記述した(下記論文)。
Yorimoto, S., Hattori, M., Kondo, M., & Shigenobu, S. (2022). Complex host/symbiont integration of a multi-partner symbiotic system in the eusocial aphid
Ceratovacuna japonica.
Iscience,
25 (12).
[注釈2] ヨーロッパの種である
Lachnus roborisの共生システムについて、極めて詳細な組織学的な研究文献、Klevenhausen (1927)を参照した(下記論文)。約100年前である当時はブフネラやセラチアは知られていなかったため、近年の細菌叢解析を含む文献も参照することで細菌種の同定を行った。
Klevenhausen F. 1927 Beiträge zur kenntnis der aphidensymbiose.
Z. Morphol. Okol. Tiere. 9, 97–165. (https://doi.org/10.1007/BF00424723)
[注釈3] ゲノム解析は、トランスオミクス解析室の協力のもと実施した。Nanopore long-read sequenceとIllumina short-read sequenceのハイブリッド戦略を用いて新規ゲノムアセンブリを行い、クリオオアブラムシの共生細菌2種:ブフネラとセラチアA系統の両方の完全ゲノムを決定した。
[注釈4] 共生細菌の置換を引き起こす要因については現在も活発に議論されており、「共生細菌のゲノム縮小進化の過程で、ある点を超えると十分な機能を果たせなくなるため、ホスト昆虫は新しい細菌を外部から取り込む必要がある」という仮説がある。この仮説と、「ホストの生態学的な変化によりパートナーの置換が駆動される」という仮説は、互いに排他的なものではなく、双方が複合的に作用していると考えられる。
【本研究に関するお問い合わせ先】
基礎生物学研究所 進化ゲノミクス研究室
助教 野崎友成
TEL: 0564-55-7672
E-mail: nozaki.t@nibb.ac.jp
【報道担当】
基礎生物学研究所 広報室
TEL: 0564-55-7628
FAX: 0564-55-7597
E-mail: press@nibb.ac.jp




