弘前大学
慶應義塾大学
自然科学研究機構 基礎生物学研究所
東邦大学
【本件のポイント】
・プラナリアの有性個体には無性個体を有性状態に誘導することのできる有性化因子が含まれている。有性化因子の投与で引き起こされる有性化過程には、有性化因子の投与がなくても有性状態を維持できるようになる特異点「有性化回避不能点」が存在している。
・有性化因子の投与で発現変動する遺伝子ライブラリを用いたトランスクリプトーム解析とRNAi法による遺伝子ノックダウン解析により、3つの有性化必須遺伝子(核内受容体をコードする遺伝子
Dr-nhr-1、転写因子をコードする遺伝子
Dr-dmd-1、
Dr-klf4l)が同定された。
・3つの有性化必須遺伝子のノックダウン個体では、有性化因子の刺激があっても、共通して精巣が分化誘導されず、有性化回避不能点を越えることができないことがわかった。これらのノックダウン個体のトランスクリプトーム解析と遺伝子ノックダウン解析により、3遺伝子の下流で働く有性化必須遺伝子として、ギャップ結合タンパク質であるイネキシンをコードする遺伝子
Dr-siriが同定された。
・プラナリアにおける有性化必須遺伝子の存在を明らかにし,本研究の成果は,今後多くの動物でみられる生殖様式転換現象の共通原理の解明に大きく貢献することが期待される。
【本件の概要】
扁形動物のプラナリアは、環境の変化に応じて分裂・再生による無性生殖と、生殖細胞を形成して他個体と交配する有性生殖を切り替えます。プラナリアは無性個体に有性個体をエサとして与えることで無性状態から有性状態に誘導(有性化)できることが以前から知られており、このことは有性個体に「有性化因子」と呼ばれる生理活性物質が含まれていることを意味しています。研究チームは有性化因子を手がかりにプラナリアの生殖様式転換の仕組みの解明に取り組みました。
本研究では、有性化因子の刺激で実験的に無性状態から有性状態への転換をうながすことができるプラナリアであるリュウキュウナミウズムシ
#1(図1)を用いて、有性化過程中の個体からRNAを抽出して、RNAシークエシング
#2を行い、遺伝子ライブラリを構築しました。そして,トランスクリプトーム解析
#3とRNAi法による遺伝子ノックダウン解析
#4により、4つの有性化必須遺伝子を同定することに成功しました。これらの有性化必須遺伝子がすべて精巣分化に関与していたことから、プラナリアの無性生殖から有性生殖への転換では、精巣分化が必須であると結論づけられました。今後、本研究の成果が手がかりとなり、多くの動物でみられる生殖様式転換現象の共通原理の解明に大きく貢献することが期待されます。プラナリアと同じ扁形動物に属する寄生性の吸虫類の多くも、陸生の巻貝を中間宿主、哺乳類を終宿主として無性世代と有性世代を転換しています。今後、吸虫類でプラナリアの有性化必須遺伝子に相当する遺伝子を明らかにすることで、吸虫類の有性化(性成熟)のメカニズムを解明することができます。そうなれば、顧みられない熱帯病とされる世界的な吸虫類による健康被害の軽減などにつながるかもしれません。
本研究成果は、2025年11月18日に国際科学誌「PLOS Genetics」に掲載されました。
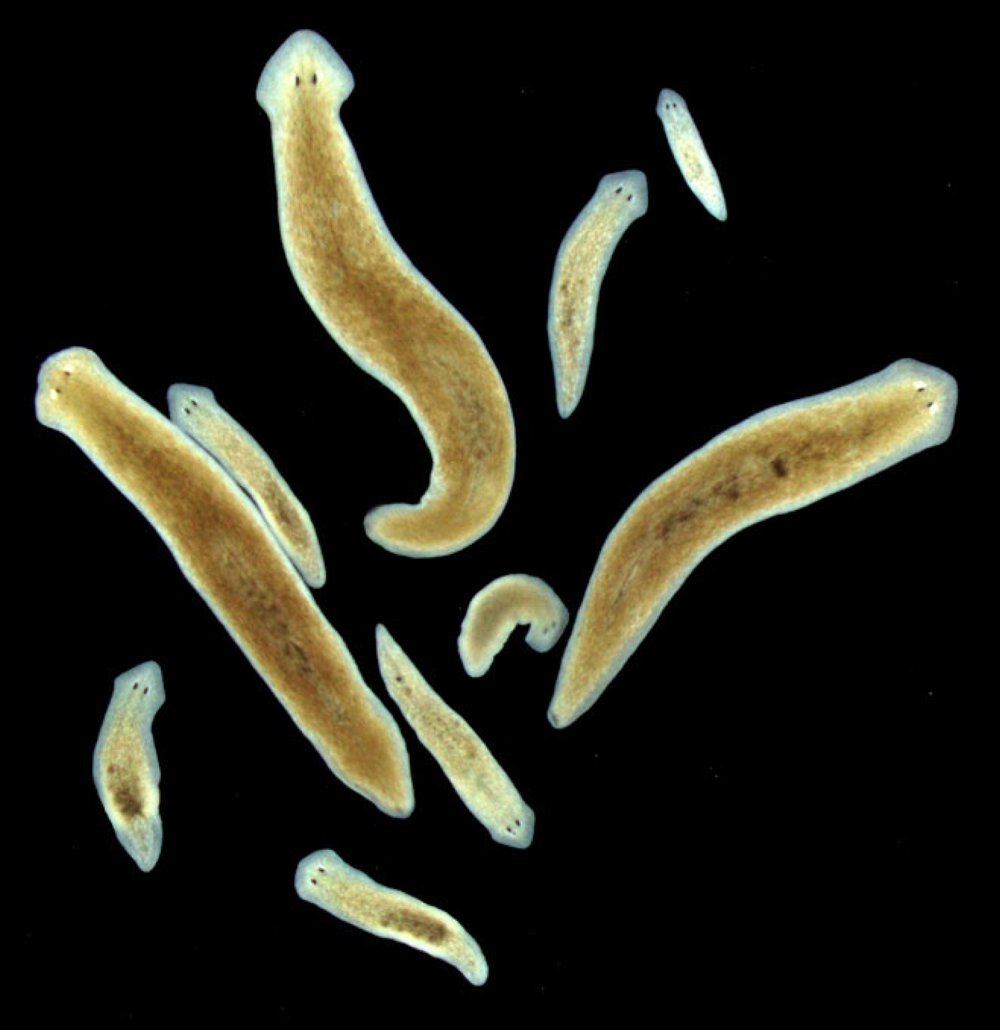 図1. リュウキュウナミウズムシの無性個体と有性個体
図1. リュウキュウナミウズムシの無性個体と有性個体
同じ遺伝子情報をもつクローン集団であるが、生殖器官をもたず分裂・再生を繰り返し増殖する無性状態(小型の個体)と生殖器官をもち交尾・産卵をすることのできる有性状態(大型の個体)がある。
【研究の背景】
「性」には男や女、♂♀、性別という意味がありますが、「同種の二個体間で遺伝子を混ぜ合わせる」という意味もあります。私たちヒトを含めた哺乳類は必ず精子と卵子が受精することで新たな生命がつくられます。まさに同種の二個体間で遺伝子を混ぜ合わせていますから、性があるわけです。一方で、「新たな生命がつくられる」ことを「生殖」といいます。これらのことをあわせて有性生殖といいます。哺乳類の生殖様式が有性生殖に限定されているために見過ごされがちですが、性を伴わないで生殖が起こる無性生殖を行う生物も多く知られています。
有性生殖と無性生殖にはそれぞれメリットとデメリットがあります。有性生殖は無性生殖に比べて遺伝的多様性をつくりやすいのですが、無性生殖に比べて繁殖スピードが遅い上に生殖コストがかかると考えられています。実は、多くの動物が無性生殖と有性生殖とを転換することができます。つまり、この生殖様式転換現象は、2つの生殖様式の「いいところ取り」をした生殖戦略ともいえます。しかし、その転換のメカニズムはほとんど明らかとなっていません。
ある種の扁形動物プラナリアは水温の変化が重要な要因となって季節的に生殖様式を転換します。無性状態から有性状態への転換は特に「有性化現象」とよばれ、プラナリアの分化多能性幹細胞「ネオブラスト」から生殖器官が分化してくることから、生殖生物学的だけでなく発生生物学的にも興味深い現象といえます。50年程前に、無性個体に有性個体を餌として与えることによって有性化が引き起こせることが報告されました。この研究によって、無性個体に生殖器官を誘導する有性個体中の化合物「有性化因子」の存在が示唆されました。弘前大学農学生命科学部の小林一也教授の研究室では、リュウキュウナミウズムシ無性クローン集団であるOH株に有性個体をエサとして与えることによる有性化の実験系(有性化系)が確立されていて(図2)、有性化機構の解明をめざす研究が行われています。
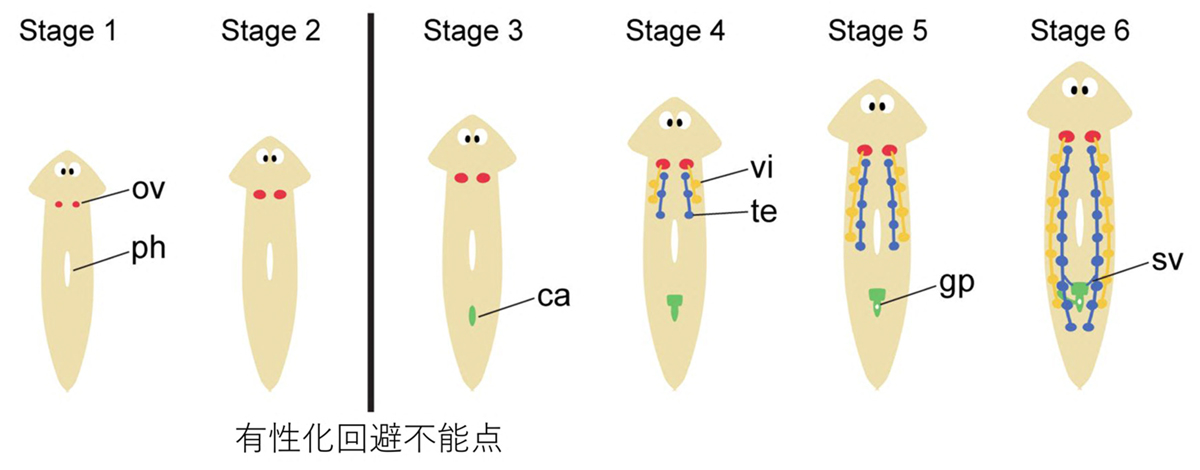 図2. リュウキュウナミウズムシの有性化過程
図2. リュウキュウナミウズムシの有性化過程
リュウキュウナミウズムシ無性クローン集団であるOH株に有性化因子を含むエサを毎日少しずつ与えることで実験的に有性化ができる。はじめに卵巣が発達し、その後精巣や卵黄腺などの生殖器官が発達してくる。その形態変化から有性化過程は6段階のステージに分けられた。ca, 交接器官; gp, 生殖孔; ph, 咽頭; ov,卵巣; sv, 貯精囊; te, 精巣; vi, 卵黄腺
有性化の過程には6つのステージがあります。ステージ3以降は有性化因子の外部投与なしでも有性化が進行する一方で、ステージ2までは有性化因子の外部投与をやめると無性状態に戻ってしまうことから、ステージ2と3の間にある特異点を「有性化回避不能点」と名付けました(図2)。
【研究の成果】
弘前大学農学生命科学部の小林一也教授のグループ、慶應義塾大学の関井清乃講師、古川亮平准教授、基礎生物研究所の重信秀治教授のグループ、および東邦大学の鹿島誠講師で構成される研究チームは、プラナリアの有性化に必須な遺伝子を単離・同定するために、無性個体に有性化因子を投与して「有性化回避不能点」に注目した遺伝子ライブラリを構築しました。そして、この遺伝子ライブラリを用いて、トランスクリプトーム解析とRNAi法による遺伝子ノックダウン解析により、4つの有性化必須遺伝子(核内受容体をコードする遺伝子
Dr-nhr-1、転写因子をコードする遺伝子
Dr-dmd-1、
Dr-klf4l、ギャップ結合タンパク質であるイネキシンをコードする遺伝子
Dr-siri)を同定しました。
プラナリアが分裂して再生することができる能力や、環境に応じて生殖器官を発達できる能力は、どのような細胞にでもなりうる未分化細胞である成体多能性幹細胞ネオブラストを持っていることと関係がありますが,無性状態ではネオブラストから生殖器官が作られることはありません。すなわち,生殖器官の分化についてはある種の「休眠状態」にあるといえます。無性個体に有性化因子を投与して「有性化回避不能点」を超えると、もはや有性化因子の投与は必要なく有性化が進行することから、有性化因子は、この「休眠状態」を打破するために必要な遺伝子、すなわち有性化必須遺伝子を「有性化回避不能点」で発現誘導していると考えられます。そこで、本研究では、有性化中の個体からRNAを抽出してRNAシークエシングを行い、遺伝子ライブラリを構築しました。そして、トランスクリプトーム解析によって有性化回避不能点直前のステージ2において発現が上昇する遺伝子の中で、ステージ6(有性個体)になった時に無性個体に比べて発現が10倍以上になっている12遺伝子を絞り込みました。
次に、これらの12遺伝子の中に有性化必須遺伝子が含まれているかどうかを確かめるために、RNAi法による遺伝子ノックダウン解析を行いました。もし、有性化必須遺伝子であれば、たとえ有性化因子が投与されていてもRNAi法により機能が低下させられていれば、有性化回避不能点を超えられないと仮定しました。その結果、核内受容体をコードする遺伝子
Dr-nhr-1、転写因子をコードする遺伝子
Dr-dmd-1、
Dr-klf4lが有性化必須遺伝子であることがわかりました。これらの3遺伝子のノックダウン個体は有性化因子の投与を停止すると、いずれも無性状態に戻りますが、有性化因子投与下では発達する生殖器官に大きな違いがありました(表1)。
Dr-nhr-1ノックダウン個体では全ての生殖器官が発達できないという結果に対して、
Dr-dmd-1や
Dr-klf4lノックダウン個体では卵巣・卵黄腺・交接器官といった生殖器官が発達します。研究チームは、いずれのノックダウン個体でも精巣だけは共通して発達してないことを見出しました。
表1. 有性化必須遺伝子のノックダウン個体での表現型
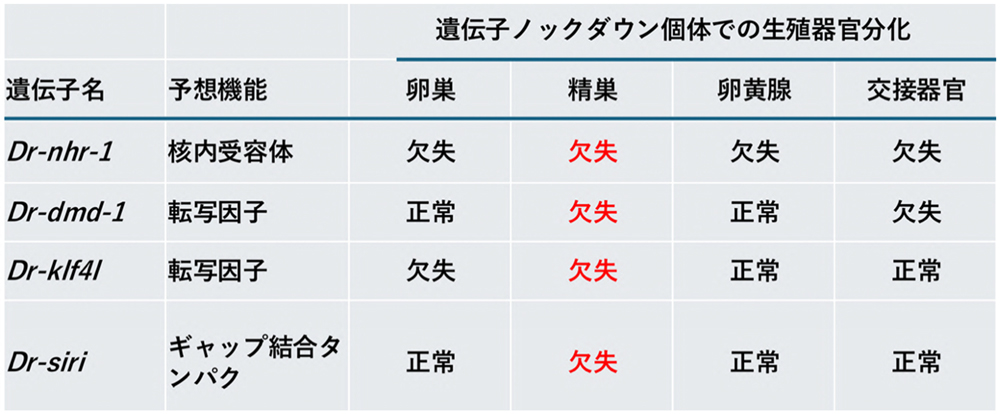
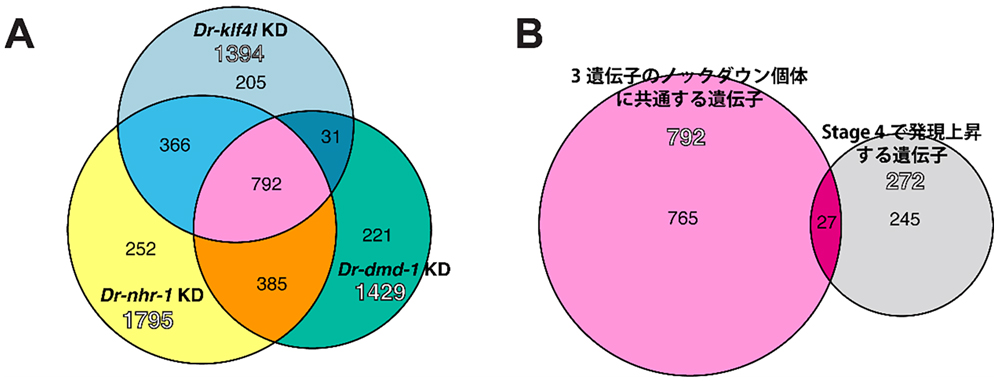 図3. Dr-nhr-1, Dr-dmd-1, Dr-klf4lの下流で精巣分化に関与する遺伝子の選定
図3. Dr-nhr-1, Dr-dmd-1, Dr-klf4lの下流で精巣分化に関与する遺伝子の選定
(A)
Dr-nhr-1,
Dr-dmd-1,
Dr-klf4lのノックダウン(KD)個体で共通して発現低下する遺伝子が792あることがわかった。(B)有性化過程ではステージ4(図2)から精巣が出現することから、ステージ4で発現上昇する272遺伝子の中で、3遺伝子のノックダウン個体で共通して発現低下する792遺伝子と重複する27遺伝子が選出された。
そこで、これらの3遺伝子のノックダウン個体からRNAを抽出して、RNAシークエシングを行い、遺伝子ライブラリを構築しました。3遺伝子のノックダウン個体で共通して発現が低下している792遺伝子中から、精巣が出現し始めるステージ4(図2)で発現上昇する272遺伝子と共通する27遺伝子を絞り込みました(図3)。私たちは、27遺伝子の中にギャップ結合タンパク質であるイネキシン遺伝子が含まれていることに気がつきました。FISH法
#5による発現解析したところ、このイネキシン遺伝子は精巣内にある精子幹細胞と体細胞であるニッチ細胞に発現していることがわかりました。さらに、有性化因子の投与下で遺伝子ノックダウン解析を行ったところ、ノックダウン個体は生殖器官の中で精巣だけが発達することができず(表1)、有性化因子の投与をやめると無性状態に戻ることがわかりました。私たちは、この遺伝子がリュウキュウナミウズムシの有性化に関わるイネキシン遺伝子であることから、その学名と英語の頭文字をとって、
Dr-siri(
Dugesia ryukyuensis-
sexual
induction
related
innexin)と名付けました。そして、私たちは、本研究で同定された4つの有性化必須遺伝子の働きを予想して次のモデルを提案しました(図4)。有性化因子の刺激でネオブラストから
Dr-nhr-1発現細胞が出現し、この細胞から
Dr-klf4lが発現する精子幹細胞の前駆細胞と
Dr-dmd-1が発現するニッチ細胞の前駆細胞が誘導されます。最終的にこれらの前駆細胞に
Dr-siriが発現することで、精子幹細胞とニッチ細胞となり精巣が分化することができると考えています。一般的に、ギャップ結合タンパク質は細胞と細胞を連絡して低分子化合物などを輸送する管として働きます。
Dr-siriが関与しているギャップ結合タンパク質は精子幹細胞とニッチ細胞をつなぎ、精巣の分化・維持に必要な低分子化合物を輸送することに働いていると予想しています。もしかしたらその中には有性化因子も含まれているかもしれません。本研究で明らかとなった4つの有性化必須遺伝子がすべて精巣分化に関与していたことから、プラナリアの無性生殖から有性生殖への転換では、精巣分化が必須であると結論づけられました。
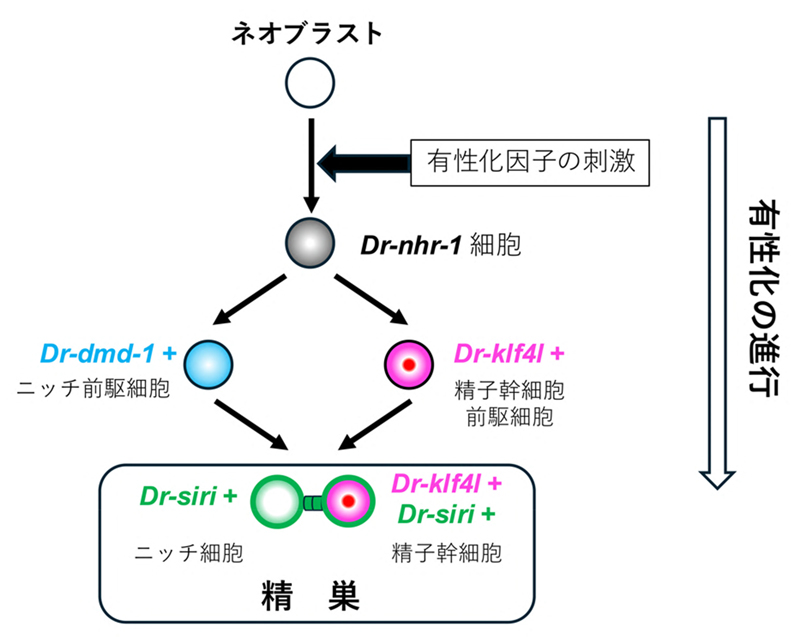 図4. 4つの有性化必須遺伝子がコードしているタンパク質の機能から予想される有性化機構
図4. 4つの有性化必須遺伝子がコードしているタンパク質の機能から予想される有性化機構
ネオブラストからまず
Dr-nhr-1発現細胞が出現し、Dr-klf4lが発現する精子幹細胞前駆細胞と
Dr-dmd-1が発現するニッチ細胞前駆細胞が誘導される。これらの前駆細胞に
Dr-siriが発現することで、精子幹細胞とニッチ細胞となり精巣が分化する。
Dr-siriが関与しているギャップ結合タンパク質は精子幹細胞とニッチ細胞をつなぎ、有性化因子など精巣の分化・維持に必要な低分子化合物を輸送することに働いていることが予想される。
【今後の展開】
プラナリアにおける有性化必須遺伝子を手がかりにした今後の研究が、多くの動物でみられる生殖様式転換現象の共通原理の解明につながると期待されます。寄生性の扁形動物である吸虫類は陸生の巻貝を中間宿主、哺乳類を終宿主として無性世代と有性世代を転換しています。研究チームはこれまでに、プラナリアで知られていた有性化因子が寄生性の扁形動物にまで広く保存されていることを明らかにしました(Sekii et al., 2023, iSCIENCE)。このことから、一見、プラナリアの生殖様式転換現象と異なる吸虫類でも、有性化因子を出発物質とするプラナリア有性化機構が進化的に受け継がれ保存されている可能性が示唆されました。今後、吸虫類でプラナリアの有性化必須遺伝子に相当する遺伝子を明らかにすることで、吸虫類の有性化(性成熟)のメカニズムを解明することができます。そうなれば、吸虫類の有性生殖を阻害するアイディアに発展させて、顧みられない熱帯病とされる世界的な吸虫類による健康被害の軽減などにつながることも夢ではないかもしれません。
【語句説明】
#1 リュウキュウナミウズムシ(
Dugesia ryukyuensis)
プラナリアは和名をウズムシとよびます。本研究では、1984年に沖縄で採集されたリュウキュウナミウズムシ(
Dugesia ryukyuensis)1個体に由来する無性クローン集団、OH株(沖縄[Okinawa]で採集して弘前[Hirosaki]で株化したことに由来する)が用いられました。
#2 RNAシークエシング
次世代シークエシング技術によって網羅的に転写産物を解読する手法。得られた配列情報から遺伝子の同定やカタログ化ができるだけでなく、遺伝子発現量の定量も可能です。
#3 トランスクリプトーム解析
生物の体の中で発現している遺伝子(mRNA)を網羅的に解析する方法。RNAシーケンシングで得られた配列情報から、どのような遺伝子がどのタイミングでどれくらいの量で発現しているかを調べることができます。
#4 RNAi法による遺伝子ノックダウン解析
二本鎖RNAと相補的な配列を持つmRNAが特異的に分解される現象を利用して、標的の遺伝子の二本鎖RNAを合成し生物に投与することで、標的遺伝子の発現を低下させて、その機能を解析できます。
#5 FISH (Fluorescence
in situ hybridization) 法
蛍光物質をつけたプローブ(標的遺伝子と相補的な塩基配列をもつ合成遺伝子)を標的遺伝子と結合させて、蛍光顕微鏡下で発現している組織や細胞を可視化する方法です。
【論文情報】
タイトル:
“Identification of an innexin required for termination of the asexual state in planarian reproductive switching”
著者名:
Nobuyoshi Kumagai
1, Michio Kuroda
1, Tosei Hanai
1, Masaki Fujita
1, Takaaki Hino
1, Shunta Yorimoto
2, Sayaka Manta
1, Shuzo Nakagawa
1, Moe Yokoyama
1, Leon Tajima
1, Riku Ito
1, Hikaru Yamada
1, Kota Miura
1, Makoto Kashima
3, Katsushi Yamaguchi
4, Shuji Shigenobu
2, 4, Ryohei Furukawa
5, Kiyono Sekii
6*, and Kazuya Kobayashi
1*(
*: 責任著者)
所属:
1. 弘前大学農学生命科学部生物学科、2. 筑波大学生存ダイナミクス研究センター、3. 東邦大学理学部生物分子科学科、4. 基礎生物学研究所、5. 慶應義塾大学自然科学研究教育センター、6. 慶應義塾大学商学部
掲載誌:
国際科学誌 PLOS Genetics
DOI:
10.1371/journal.pgen.1011944
本研究は科学研究費補助事業、新学術領域研究「動物における配偶子産生システムの制御」(課題番号16H01249)、科学研究費補助事業、新学術領域研究「配偶子インテグリティの構築」(課題番号19H05236)、科学研究費補助事業、基盤研究(B)(課題番号19H03256)、科学研究費補助事業、基盤研究(B)(課題番号25K02303)、基礎生物学研究所共同利用研究(14-736)によって支援されました。
【取材に関するお問い合わせ先】
(所属) 弘前大学農学生命科学部生物学科
(役職・氏名) 教授・小林一也
(電話) 0172-39-3587
(E-mail) kobkyram@hirosaki-u.ac.jp
(所属) 慶應義塾大学商学部
(役職・氏名) 専任講師・関井清乃
(電話・FAX) 045-566-1337
(E-mail) kiyono.sekii@keio.jp
【報道に関するお問い合わせ先】
(所属) 基礎生物学研究所 広報室
(電話) 0564-55-7628 FAX:0564-55-7597
(E-mail) press@nibb.ac.jp
(所属) 学校法人東邦大学 法人本部経営企画部
(電話) 03-5763-6583 FAX:03-3768-0660
(E-mail) press@toho-u.ac.jp




