自然科学研究機構 基礎生物学研究所
東京大学
お茶の水女子大学
立命館大学
ダイズに代表されるように、種子植物は細胞内の液胞と呼ばれる細胞小器官に大量のタンパク質を貯蔵し、それがヒトや家畜の主要なタンパク源になっています。このような液胞の機能は植物以外にはありません。植物は大量のタンパク質を液胞に輸送する仕組みを、進化の過程で独自に獲得してきたのです。では、他の生物にはないこの独自の物質輸送経路は、どのようにして誕生したのでしょうか。
東京大学大学院 農学生命科学研究科の藤本優准教授、基礎生物学研究所 細胞動態研究部門の海老根一生助教(研究当時/現 埼玉大学)、金澤建彦助教、南野尚紀特任助教(研究当時/現 福岡大学)、上田貴志教授(責任著者)、理化学研究所 光量子工学研究センターの清水優太朗大学院リサーチ・アソシエイト(研究当時/現 ボルドー大学)と中野明彦副センター長(研究当時/現 東京科学大学)、お茶の水女子大学 ヒューマンライフサイエンス研究所の伊藤容子特任助教と基幹研究院の植村知博教授、立命館大学 生命科学部の深尾陽一朗教授による共同研究グループは、植物独自の液胞への物質輸送経路が、VAMP7という膜どうしの融合を司るタンパク質に生じた段階的な機能変化によって開拓された過程を明らかにしました。
具体的には、植物の進化の過程でVAMP7に起きたアミノ酸挿入配列の出現とその配列の酸性化、さらに、それに伴う積み荷選別タンパク質複合体AP-4との結合能の獲得とその強化が、種子植物に特有の液胞輸送経路の成立を導く鍵となったことを突き止めました。
本研究は、細胞内の物質輸送システムを例として、植物が独自の細胞機能を発達させてきた過程を分子レベルで示したもので、その成果は2025年5月13日付けで米国の国際学術誌「Current Biology」に掲載されました。
【研究の背景】
真核細胞では、小胞や小管を介した物質輸送システム「膜交通(メンブレントラフィック)」により、様々な細胞小器官の間で物質のやり取りを行っています。このシステムを機能させるための基本要素はすべての真核生物に共通して備わっていますが、そこから派生する輸送経路やその調節のしくみは、植物や動物、酵母といった、それぞれの生物の系統で独自の進化を遂げてきました。
「液胞」は植物細胞を特徴付ける細胞小器官であり、物質の貯蔵や分解、空間充填、細胞の恒常性維持などの植物の生存や成長に不可欠な役割を果たしています。膜交通システムに関する植物の特徴として、液胞への輸送経路が多様化し、複雑なネットワークを形成していることが挙げられます。このネットワーク中を行き来する小胞には、VAMP7と呼ばれる膜交通タンパク質が積み込まれており、これが小胞とその輸送先の細胞小器官との間の膜融合を実行しています。
植物のVAMP7にはVAMP71とVAMP72という二種類のタイプがあり、基本的に、VAMP71は液胞での膜融合で、VAMP72は細胞膜への輸送経路(以下、分泌経路)でそれぞれ機能することが分かっています。上田教授らの研究グループではこれらに加え、シロイヌナズナをはじめとした種子植物にVAMP727という液胞への輸送経路(以下、液胞輸送経路)で機能する特殊なVAMP72が存在していることを突き止めていました。このVAMP727は、N末端側のロンジンドメインという領域に、他のVAMP72にはない、約20アミノ酸からなる酸性アミノ酸に富んだ配列が挿入されているという特徴を持ちます(図1)。
このVAMP727が機能する輸送経路については、種子貯蔵タンパク質の液胞への大量輸送に極めて重要であることが判明しています。しかし、VAMP727やそれに依存する液胞輸送経路が、植物の進化の過程で、いつどのようにして生まれたのか、そしてその背後にどのような分子メカニズムの変化があったのか、という点は不明でした。
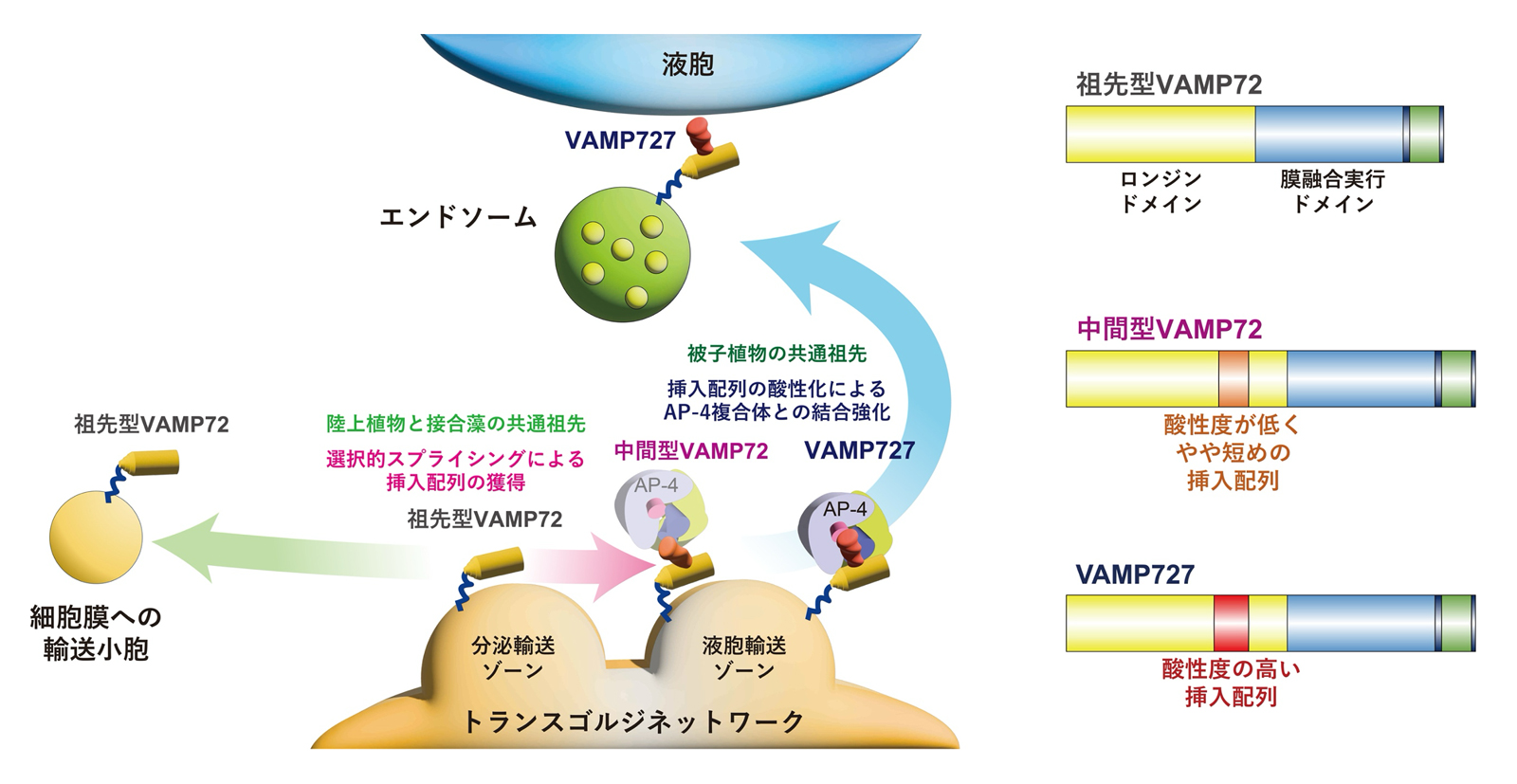
図1 植物におけるVAMP72の構造変化と液胞輸送経路の進化モデル
【研究の成果】
研究グループは本研究において、VAMP727に特有の酸性アミノ酸に富んだ挿入配列(以下、酸性挿入配列)が、液胞輸送経路において積み荷の選別を担うAP-4複合体との結合に必要であることを明らかにしました。さらに、この配列の起源が、陸上植物とそれに最も近縁な生物とされる接合藻類との共通祖先において
VAMP72遺伝子に生じた選択的スプライシングにあることを明らかにしました。AP-4複合体との結合を通じて、もともと分泌経路で機能していたVAMP72が、液胞輸送経路でも機能するようになり、この変化こそが、種子植物におけるVAMP727の誕生と、新たな液胞輸送経路の成立を導いた決定的な要因であったと考えられます。本研究の詳しい内容は以下の通りです。
VAMP727に特有の酸性挿入配列の機能
VAMP727はロンジンドメインの酸性挿入配列の有無以外は、分泌経路で働く通常のVAMP72と極めて似通った構造を持ちます。このことから、酸性挿入配列が入ることにより、VAMP727の機能が分泌経路から液胞輸送経路へと転換したのではないかと考えました。そこで、この酸性挿入配列の機能を検証するために、その有無や酸性度の違いがVAMP727の局在や機能に与える影響を解析しました。その結果、配列を欠失もしくは酸性度を低下させると、VAMP727はエンドソーム(液胞輸送経路の中継点)からトランスゴルジネットワーク(液胞輸送経路と分泌経路の分岐点)へと局在を変え、さらに液胞膜における膜融合活性が弱まる一方、細胞膜における膜融合活性が強まることが分かりました(図2)。これにより、酸性挿入配列はVAMP727が液胞輸送経路で機能するために不可欠であり、この配列の欠失は通常のVAMP72への機能の先祖返りを引き起こすことが示されました。
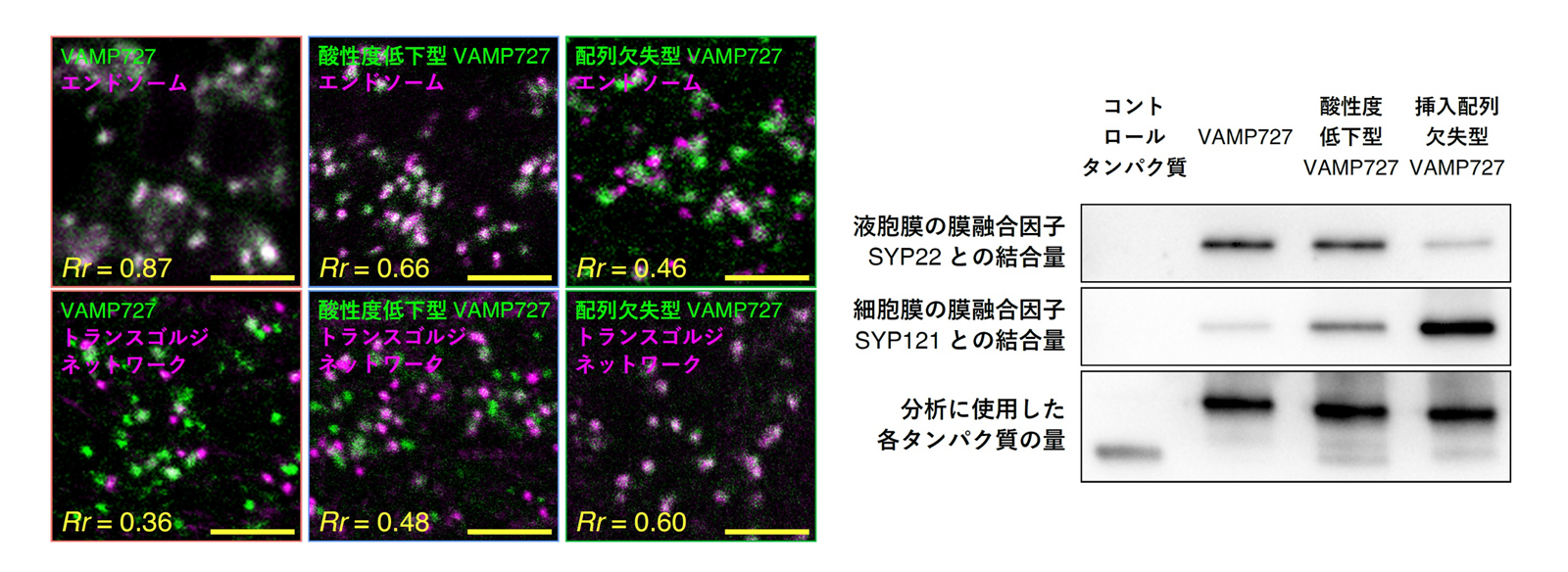
図2 酸性挿入配列の変化がVAMP727の局在や機能に与える影響
左図は、シロイヌナズナの根の細胞において、酸性挿入配列を改変したVAMP727の細胞内局在を、エンドソームやトランスゴルジネットワークの蛍光マーカーと比較した結果を示しています。右図は、各改変型VAMP727の膜融合活性、すなわち、液胞膜の膜融合因子SYP22および細胞膜の膜融合因子SYP121との結合量を比較した結果で、バンドの濃さがそれぞれの結合量を示しています。酸性挿入配列の酸性度を低下させたり、配列自体を欠失させたりすると、VAMP727のエンドソームへの局在や液胞膜での膜融合活性は低下する一方で、トランスゴルジネットワークへの局在や細胞膜での膜融合活性は上昇しました。各スケールバーは5 µmを示しています。
酸性挿入配列を介したAP-4複合体との結合とその機能的意義
次に、VAMP727の酸性挿入配列の機能をより詳しく解明するため、シロイヌナズナを用いてVAMP727と結合するタンパク質を探索し、その結合と酸性挿入配列との関連を調べました。その結果、トランスゴルジネットワークにおいて液胞方向へ向かう輸送小胞への積み荷の積み込みを担うAP-4複合体がVAMP727と結合することが分かりました。この結合は、酸性挿入配列の酸性度を低下させると弱まり、配列を欠失させると消失することから、VAMP727の酸性挿入配列がAP-4複合体との結合に不可欠であることが示されました。さらに、AP-4の機能を失ったシロイヌナズナ変異体では、VAMP727が通常のVAMP72と同様にトランスゴルジネットワークに局在するようになり、液胞膜での膜融合活性が低下する一方で、細胞膜での活性が増加することが明らかとなりました。さらに、トランスゴルジネットワークでのVAMP727の輸送過程を超解像ライブイメージング顕微鏡(SCLIM)を用いて観察した結果、AP-4複合体がトランスゴルジネットワークにおいてVAMP727をエンドソームへと輸送するのに必要であることが確認されました(図3)。これらの結果から、VAMP727が液胞輸送経路で適切に機能するためにはAP-4の働きが必須であり、進化の過程で酸性挿入配列を獲得したことで、VAMP727が液胞輸送経路における新たな機能を獲得したことが明らかになりました。
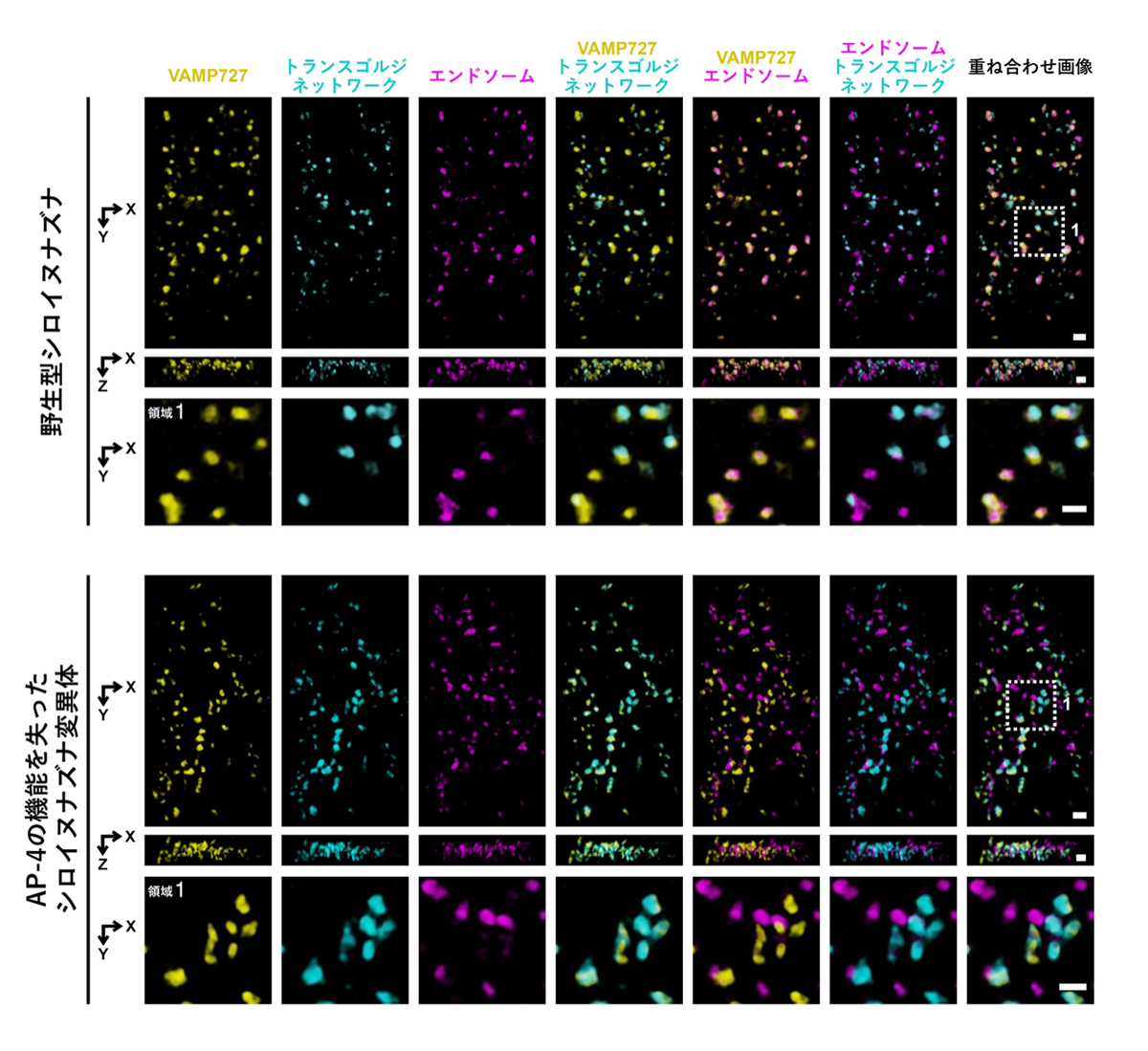
図3 AP-4複合体の機能が失われることによって生じるVAMP727の細胞内局在の変化
超解像ライブイメージング顕微鏡(SCLIM)を用いて、VAMP727の細胞内局在をエンドソームおよびトランスゴルジネットワークの蛍光マーカーと、詳しく比較した画像を示しています。上段は野生型シロイヌナズナ、下段はAP-4複合体の機能が失われた変異体の根の細胞の画像です。AP-4の機能が失われることにより、VAMP727の局在がエンドソームからトランスゴルジネットワークへと変化することが分かりました。各スケールバーは1 µmを示しています。
酸性挿入配列はどこから来たのか?
酸性挿入配列がどのように生じたのかを解明するために、様々な植物からVAMP727と類似したVAMP72の配列を収集し、それらを比較しました。その結果、酸性度の高い挿入配列を持つVAMP727とよく似た分子は被子植物のみに認められました。一方で、意外なことに酸性度が低く短めの配列が挿入されたタイプのものが、裸子植物やコケ植物のゼニゴケ、さらに陸上植物の姉妹系統である接合藻の一部に見つかりました(図4)。そこでゼニゴケのVAMP72をさらに詳しく調べたところ、挿入配列の有無だけが異なる二つのVAMP72が存在しており、これらがひとつの遺伝子からの選択的スプライシングにより生じることが判明しました。このことにより、酸性挿入配列の起源が、陸上植物と接合藻の共通祖先で初めておこった選択的スプライシングにあることが示されました。
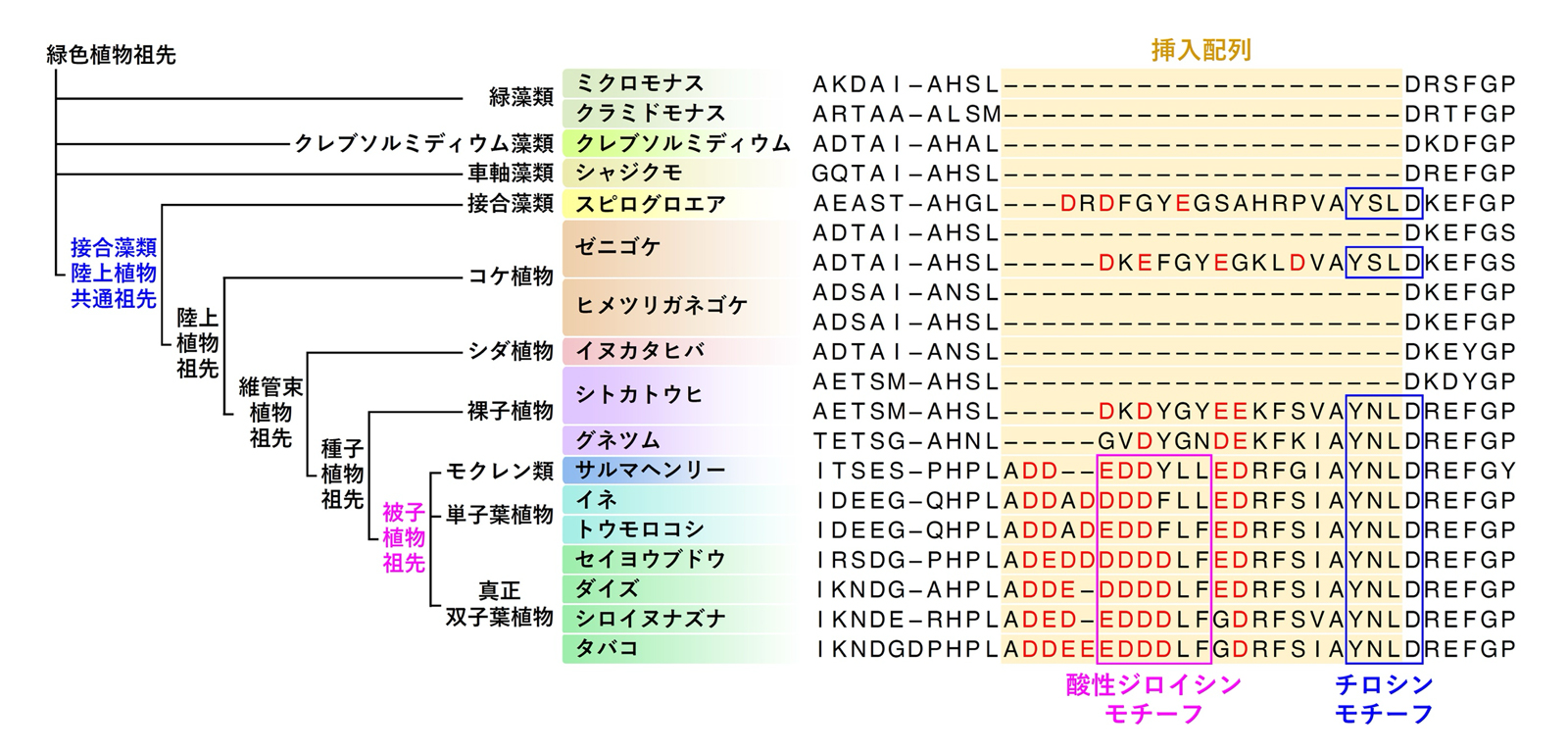
図4 VAMP72に生じた挿入配列の進化とその特徴
図の左側には、緑藻類から被子植物までの主要な植物系統を示した系統樹を、右側には、それぞれの系統に存在するVAMP727やそれに近縁のVAMP72の挿入配列付近のアミノ酸配列を示しています。
VAMP72の段階的な新機能化とその分子メカニズム
もともと分泌経路で機能していたVAMP72が、液胞輸送経路で機能するようになった過程を推定するため、緑藻から被子植物にいたる様々な植物のVAMP727やVAMP72の配列や機能を詳細に比較しました。その結果、挿入配列を持たない祖先型なタイプは分泌経路で、酸性度の高い挿入配列を持つタイプは液胞輸送経路で機能していることが分かりました。一方、酸性度が低くやや短めの配列を持つ中間型(ゼニゴケやシトカトウヒのスプライシングバリアントなど)は、両者の中間的な性質を示しました。
シロイヌナズナVAMP727を始めとする酸性挿入配列には、AP-4複合体などのクラスリンアダプターと直接結合する「チロシンモチーフ」や「酸性ジロイシンモチーフ」が含まれており、VAMP72の機能転換において重要な役割を果たしたと考えられます。一方、短めの挿入配列を持つ中間型VAMP72には「チロシンモチーフ」は含まれますが、「酸性ジロイシンモチーフ」はありませんでした(図4)。これらの結果から、VAMP72はまずチロシンモチーフを含む挿入配列を獲得することで部分的な機能転換がおこり、それが酸性化するとともに酸性ジロイシンモチーフが生じることでAP-4複合体との結合が強化され、液胞輸送経路で機能するVAMP727へと進化した可能性が示唆されます。このVAMP727の二段階の機能転換を経て、植物に新たな液胞輸送経路が誕生し、これにより液胞へのタンパク質の大量輸送が可能になったと考えられるのです。
【今後の展望】
今回の研究では、種子植物のVAMP727に特有の酸性挿入配列の機能と進化的起源を解明することを通じ、既存の膜交通タンパク質VAMP72に生じた段階的な機能転換が、新たな液胞輸送経路を開拓する原動力となったことが示されました。
本研究で得られた知見は、将来的には有用物質の貯蔵や効率的生産、植物のストレス耐性の向上などを可能にする、膜交通経路の制御技術の確立に向けた基盤となるものと期待されます。
【発表雑誌】
雑誌名 Current Biology
掲載日 2025年5月13日
論文タイトル: Neofunctionalization of VAMP7 opened up a plant-unique vacuolar transport pathway
著者: Masaru Fujimoto, Yutaro Shimizu, Yoko Ito, Kazuo Ebine, Naoki Minamino, Takehiko Kanazawa, Yoichiro Fukao, Akihiko Nakano, Tomohiro Uemura, Takashi Ueda
DOI:
https://doi.org/10.1016/j.cub.2025.04.062
【研究グループ】
本研究は、基礎生物学研究所 細胞動態研究部門、東京大学大学院 農学生命科学研究科、理化学研究所 光量子工学研究センター、お茶の水女子大学 基幹研究院及びヒューマンライフサイエンス研究所、立命館大学 生命科学部が参加した共同研究チームにより実施されました。
【研究サポート】
本研究は、科学研究費助成事業(19H05670, 19H05675, 21H02515, 24K02050, 23KF0078, 21K06222, 17H05019, 24H00509, 22K19327, 22H02643, 10J08869, JP21J00910)、生命創成探究センター(ExCELLS)特別共同研究プログラム(No. 23-S3 and 24-S2)、三菱財団、山田科学振興財団、および、旭硝子財団の支援を受けて行われました。
【本研究に関するお問い合わせ先】
基礎生物学研究所 細胞動態研究部門
教授 上田貴志
TEL: 0564-55-7530
E-mail: tueda@nibb.ac.jp
【報道担当】
基礎生物学研究所 広報室
TEL: 0564-55-7628
FAX: 0564-55-7597
E-mail: press@nibb.ac.jp
東京大学大学院農学生命科学研究科・農学部
総務課総務チーム 総務・広報情報担当(広報情報担当)
お茶の水女子大学 広報・ダイバーシティ推進課
立命館大学 総合企画部広報課




