基礎生物学研究所



2016.05.20
国立大学法人 筑波大学
大学共同利用機関法人 自然科学研究機構 基礎生物学研究所
国立大学法人筑波大学生命環境系 和田洋教授、同大学院 岡田和訓博士(現在は基礎生物学研究所 研究員)、岡崎統合バイオサイエンスセンター/基礎生物学研究所 高田慎治教授らは、メダカの鰓がかたち作られる初期段階で、鰓のもとになるくり返し構造が作られるためにpax1という遺伝子が鍵となる役割を果たすことを発見しました。
生き物の体のかたちができあがる途中段階では、同じようなかたちのモジュール構造がくり返し作り出され、その後の発生過程で組織や器官のかたちに反映されることが知られています。たとえば、私たち脊椎動物の背骨は、椎骨が何個も数珠のように繰り返し連なっていることがわかります。これは、体節と呼ばれるくり返し構造が発達してできたものです。体節のくり返し構造が形成される仕組みについては研究が進んでおり、基本的な分子機構が解明されつつあります。脊椎動物では、体節の他に鰓にもくり返し構造が見られます。一般的な硬骨魚では、鰓は4枚並んでいますが、このような鰓のくり返し構造がどのような仕組みでできあがるかについてはこれまでわかっていませんでした。
本研究ではメダカのpax1という遺伝子注1に注目し、この遺伝子の発現が表すくり返しのパターンが、鰓のもとになるくり返し構造を作るための基準になっていることを明らかにしました。さらに、ゲノム編集という方法を用いてメダカのpax1の機能をなくした突然変異体を作成したところ、鰓が形成されないことがわかりました。最も重要な発見は、pax1の突然変異体では、pax1自身のくり返しパターンでの発現が見られなくなったことです。この研究結果は、鰓のくり返し構造の形成にpax1が中心的な役割を果たしていることを示しています。今後はこの成果をもとに、pax1がどのような仕組みでくり返しのパターンを生み出しているかを解明することで、生物がくり返し構造を作り出していく仕組みについて理解が進むことが期待されます。
本研究の成果は英国の国際発生生物学専門誌「Development」電子版で、5月15日付で公開されました。
本研究は日本学術振興会の科学研究費補助金のもとで行われました。
【研究の背景】
生物は進化の様々な局面でくり返し構造を生み出してきました。たとえば、ゴカイやムカデの体には顕著なくり返し構造が見られます。私たちの体の背骨を見ても、多数の椎骨が連結していることからわかるように、脊椎動物の体にもくり返し構造があります。これらのくり返し構造は、異なる系統で独立に進化したと考えられています。脊椎動物のからだをよく見ると、背骨の他に鰓にもくり返し構造があります(図1)。たとえば、サメを横から見ると鰓穴が並んでいるのが見えます。一般的な硬骨魚も鰓蓋を開いてみると鰓が4枚並んでいるのがわかります。背骨のもとになる体節と呼ばれるくり返し構造の作られ方については理解が進んでいますが、鰓のくり返し構造の作られ方についてはよくわかっていませんでした。
【研究内容と成果】
本研究では、メダカのpax1という遺伝子に着目して研究しました。pax1がマウスやニワトリで鰓のもとにあたる細胞で発現していることは先行研究により知られていました(マウスやニワトリは陸上に上がったため鰓は失っていますが、胚の時期には鰓のもとになるくり返し構造が出現します)。しかしマウスのpax1突然変異体では、鰓状のくり返し構造の形成がほぼ正常に進むことから、pax1が鰓のくり返し構造の形成に関与することは予想されていませんでした。
まず、私たちはメダカの鰓形成におけるpax1の発現を詳細に解析ました。pax1が鰓で発現することはわかっていましたが、pax1の発現がいつ始まるのか、その発現が鰓のくり返し構造を形作ることと関係があるのか、ということに関しては詳しく調べられていませんでした。発生の過程で、鰓のもとになる構造は前から後ろに一個ずつ、くり返し形成されます。そこで、pax1の発現する場所を、鰓のもとになる構造が作られ始める前の時期、作られ始めた時期、作られ終わった時期、そして次の構造の作られ始めの時期に、という具合に順を追って調べました。すると、pax1の発現は鰓のもとになる構造ができ始めるよりも早いタイミングで始まり、その場所から鰓のもとになる構造ができていくことがわかりました。この現象がくり返し見られたことから、pax1の繰り返しの発現が鰓のもとになる構造が繰り返し形成される合図になっていると考えられました。
そこで、東京工業大学の研究グループと共同し、ゲノム編集技術によってpax1の突然変異体を作成して鰓の形成を解析しました。すると、pax1の突然変異体では鰓の形成が妨げられることがわかりました(図2)。そして、突然変異体での鰓形成の異常は、メダカが卵から孵化する前の発生初期の段階で、鰓のもとになるくり返し構造を作れなくなったためであることもわかりました(図3)。先に行ったpax1発現パターンの解析結果と合わせて考えると、pax1が発現して機能する細胞と発現しない細胞がくり返し現れることが、鰓のくり返し構造の基準になっていることがわかりました。
では、pax1の発現パターンはくり返しパターンの最初の合図なのでしょうか?それとも、pax1の発現よりも先にできていたくり返しパターンを写し取っているだけなのでしょうか?私たちはさらに解析を進め、pax1の突然変異体において、pax1のmRNAの発現を調べました。pax1の突然変異体では正常に機能するPax1タンパク質を合成することはできませんが、mRNAの合成は起こります。つまり、pax1のくり返しパターンの発現が別の遺伝子の作ったパターンの丸写しであるならば、pax1の突然変異体でもpax1 mRNAのくり返しパターンは観察されるはずです。しかし、実際にはpax1 mRNAの発現が一様なパターンになり、本来のくり返しパターンが失われていることがわかりました(図4)。この結果は、pax1の機能が自分自身のくり返しパターンでの発現に、さらにはくり返し構造の形成に必須であることを示しています(図5)。
私たちヒトを含め、陸上に上がった脊椎動物は鰓を持っていません。しかし、私たちも胚発生の時期には鰓のもとになるくり返し構造を持っています。そして、それらは鰓を作らない代わりに私たちの生存に必要な様々な器官を作り出しています。これまでに研究された全ての脊椎動物で、このくり返し構造の形態、形成される位置、そしてそこで発現する遺伝子の種類までもが非常によく似ていたことから、脊椎動物はどの種であっても一旦同じようにくり返し構造を作り、その後で各種の生活様式に合わせた器官へと作り変えているのであろうと考えられてきました。しかし意外なことに、今回の結果はくり返し構造の形成機構そのものにメダカとマウスでは決定的な違いがあることを示しています。つまり、私たちの祖先が水中から陸上の環境に適応していった進化の過程で、これまで考えられていたよりも抜本的な体の作り方の改変があった可能性が出てきました。本研究によってメダカの鰓の作られ方が解明されたことで、全ての脊椎動物に共通した構造の裏側に隠されていた、予想外な進化の側面が見えてきたのです。
【今後の展開】
今後、pax1のどのような機能が、くり返し構造を作り出していくかをさらに詳細に解析することで、鰓におけるくり返し構造の形成機構を解析できることが期待されます。
生物は進化の歴史のさまざまな局面でくり返し構造を進化させてきました。このような進化の戦略がどのような仕組みで成し遂げられてきたのか、多種多様な生物を生み出した仕組みは一様なのか、あるいはもっと臨機応変だったのか、動物進化の壮大な歴史の一端の解明を目指します。
参考図
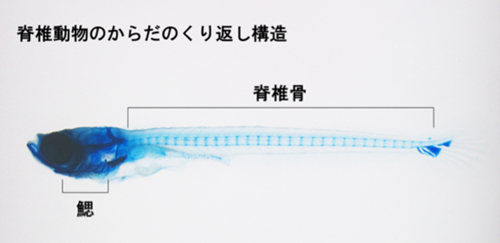
図1;メダカ幼魚の軟骨組織の染色写真。
頭には鰓、体幹には脊椎骨という二つのくり返し構造があることがわかる。
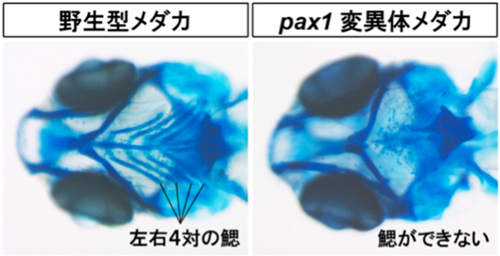
図2;野生型メダカとpax1突然変異体の鰓の骨格。
写真のように腹側から観察すると野生型メダカでは左右4対の鰓が並んでいることがよくわかる。一方、pax1変異体メダカでは鰓の骨格が形成されていない。
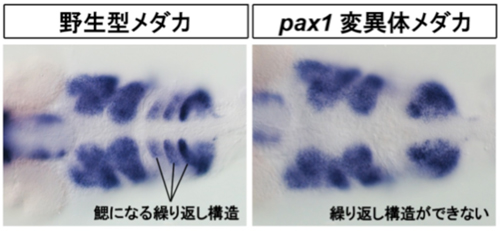
図3;胚発生の時期に頭部に形成されるくり返し構造を染色した写真。
胚時期に現れるくり返し構造は発生が進むと鰓になる。pax1突然変異体では野生型に見られるくり返し構造が作られない。
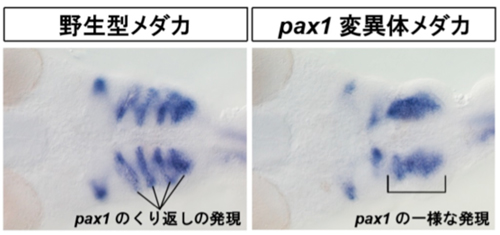
図4;メダカ胚の頭部におけるpax1 mRNAの発現パターン。
野生型で見られるpax1発現のくり返しパターンを、pax1変異体は作り出すことが出来ない。
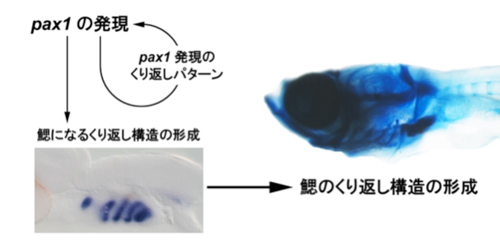
図5;メダカの鰓の形成におけるpax1の役割。
pax1が発現することでpax1自身の発現のくり返しパターンを作り出す。これによってpax1が発現する細胞としない細胞がくり返し頭部に並ぶ。pax1が発現した細胞が鰓を作るので、メダカはくり返し構造として鰓を形成することができる。
用語解説
注1) pax1遺伝子:脊椎動物では背骨を形成する細胞が分化する際に必須な役割を果たすことがわかっている。鰓のもとになる細胞で発現することもわかっていたが、マウスの突然変異体では鰓のもとになるくり返し構造にはほとんど異常がないためその機能はよくわかっていなかった。
掲載論文
【題 名】 Reiterative expression of pax1 directs pharyngeal pouch segmentation in medaka
(和文タイトル;pax1のくり返しの発現が鰓のくり返し構造の発生の指揮をとる)
【著者名】 Kazunori Okada, Keiji Inohaya, Takeshi Mise, Akira Kudo, Shinji Takada, and Hiroshi Wada
【掲載誌】 Development doi:10.1242/dev.130039
【問合わせ先】
和田 洋 (わだ ひろし)
筑波大学 生物環境系 教授
〒305-8572 茨城県つくば市天王台1-1-1
E-mail: hwada@biol.tsukuba.ac.jp
Tel: 029-853-4671
高田 慎治 (たかだ しんじ)
岡崎統合バイオサイエンスセンター / 基礎生物学研究所 分子発生学研究部門 教授
〒444-8787 愛知県岡崎市明大寺町東山5-1
E-mail: stakada@nibb.ac.jp
Tel: 0564-59-5241
岡田 和訓 (おかだ かずのり)
基礎生物学研究所 分子発生学研究部門 研究員
〒444-8787 愛知県岡崎市明大寺町東山5-1
E-mail: okada@nibb.ac.jp
Tel: 0564-59-5243

