多くのほ乳類において、受精後発生を進める胚が着床前の段階で一時的に発生を休止することが知られており、これは発生休止(遅延着床とも呼ばれる)と呼ばれる現象である。母親が置かれた環境が整うと、発生を再開する。
基礎生物学研究所 初期発生研究部門の亀水千鶴研究員、藤森俊彦教授の研究グループはマウス胚が発生休止に入る過程、休止中からの発生を再開する過程を細胞レベルで観察した。発生休止中の胚においても細胞の分化状態は維持されたままであるが、発生休止過程が複数のステップに分離できること、細胞周期の停止が胚の領域や分化形質によって異なることを明らかにした。また、発生休止からの再開においても胚の領域によって進み方が異なることから、発生休止を制御する機構が胚の領域によって異なることが示唆された。本成果はBiology of Reproduction誌に掲載された。
【研究の背景】
ほ乳類の体作りは卵と精子が卵管で出会うことで始まり、受精卵は細胞分裂を繰り返しやがて子宮に着床し、胚発生が進む。100種以上のほ乳類において、受精した卵から発生を始めた後に子宮に着床する前の段階(胚盤胞)で発生を一旦止める現象が知られており、発生休止と呼ばれている(着床が遅れることから遅延着床とも呼ばれる)。種によっては、数ヶ月間発生を停止する。一旦発生を休止した胚は、その後子宮内の環境が整うと着床し発生を再開する。発生休止は季節や飢餓状態、授乳など、母親が置かれた環境に応じて一旦発生を止め、母親の環境が整うと胚は発生を再開する生存戦略の一つであると考えられている。例えば、野生のクマは、春から夏の間に受精した卵が発生休止し、母親が冬眠する前に着床し、冬眠中に出産することが知られている。このように、発生休止という現象は、ほ乳類胚では一般的な生存戦略であると考えられるが、発生休止が誘導され、胚盤胞の段階で発生を休止しつづける分子機構については未解明である。
研究に用いられているマウス(ハツカネズミ)でも、母親が授乳中に交尾した場合には、受精が成立した後に、着床前の段階で胚の発生が休止することが知られている。胚は卵管で受精した後に卵管で発生を開始し、子宮に移動した胚盤胞の段階で着床する。実験的にも交尾後3日目に卵巣を除去し、プロジェステロンの投与を繰り返し行うことで、発生休止を誘導することができる。この系を利用して、マウスの発生休止胚を経時的に観察することによって、胚盤胞がどのようにして発生休止状態に入り、発生休止から復帰するかを明らかにすることを目指した。発生休止誘導後1週間にわたって経過を解析した。
まず、発生休止が確実に誘導できているかを確認するために、休止誘導後の各段階において胚盤胞を回収し、2.5日目の偽妊娠マウスの子宮に移植した。全ての段階に回収した胚は、移植後着床し胚発生が問題なく進められたことから、発生休止の誘導が確実に行われていた。休止期間が長くなると、移植後の胚発生の遅延がみられたことから、発生再開までに時間を要することが示唆された。
【研究の成果】
受精から胚盤胞の早い時期までの間、卵(胚)は、透明帯と呼ばれる膜に包まれた状態で発生が進む。着床に際して胚は透明帯から飛び出し(ハッチし)、直接子宮の上皮細胞と接着出来る状態に入る。子宮の中での胚を観察すると、休止中の胚は透明帯からハッチしているが胚は子宮上皮には直接接触しておらず、子宮上皮の上皮構造も維持されており脱落膜反応も見られていないことから、着床時に見られる子宮の反応が無いことが示唆された。各段階の発生休止胚を子宮から取り出し観察すると、着床前の胚盤胞に比較してサイズが大きく、胚−非胚軸(Em-Ab軸)に沿って伸長しており、通常の着床前胚とは形態的に異なることが明らかになった。
発生休止中の胚について、細胞分化、細胞数、細胞周期に着目して解析を進めた。着床直前の胚においては、栄養外胚葉、原始内胚葉とエピブラストの3種類の細胞が見られる。それぞれのマーカーであるCdx2, Gata4, Nanogの発現を抗体染色した。それらは発生休止の全ての時期において発現しており、原始内胚葉とエピブラストは混在せず層構造を形成していたことから、正常発生における着床直前の状態で分化状態は維持されていることが示唆された。
次に、発生休止中の胚の中での細胞数の変化を解析した。休止誘導して3日目までは緩やかに細胞数の増加が見られ、その後約200個で細胞数が一定となった。分化マーカーと核染色とを比較したところ、分化形質と細胞数の増加に関しては明確な関連は見られなかった。細胞数の変化が細胞周期の変化による物か調べるために、細胞周期を可視化するためのR26-Fuscci2マウスを用いて、休止胚を回収後ただちに観察した。非休止胚の胚盤胞はほぼ全ての細胞では緑色の蛍光がみられ、G1あるいはG0期は短い。一方で発生休止胚については、内部細胞塊から離れた(非胚領域の)栄養外胚葉ではほとんどの胚で赤色の蛍光がみられたことから、G1あるいはG0期であった。休止後1日目には内部細胞塊だけでなく、胚側の栄養外胚葉でも緑色の蛍光が見られる細胞が存在していたが、休止時期が長くなるに従って、栄養外胚葉は赤色の蛍光のみとなり、6日目にはほぼ全ての細胞が赤色となった。このことから、発生休止誘導後、全ての細胞が同時に細胞周期を止めるのではなく、非胚側の栄養外胚葉から細胞周期を止め、やがて胚側の栄養外胚葉、最後に内部細胞塊領域で細胞周期が止まることが示唆された。別の細胞周期マーカーであるKi67抗体を用いて各段階の発生休止胚を染色した場合にも、細胞周期の止まり方については、Fucci2を用いた観察と同じ傾向がみられた。休止誘導後6日目にはわずかにKi67陽性細胞が存在していたが、休止誘導後7日目には全ての細胞がKi67陰性であった。この事から、細胞周期は非胚領域から緩やかに停止し、休止誘導後7日目には全ての細胞がG0期にあることが示唆された。
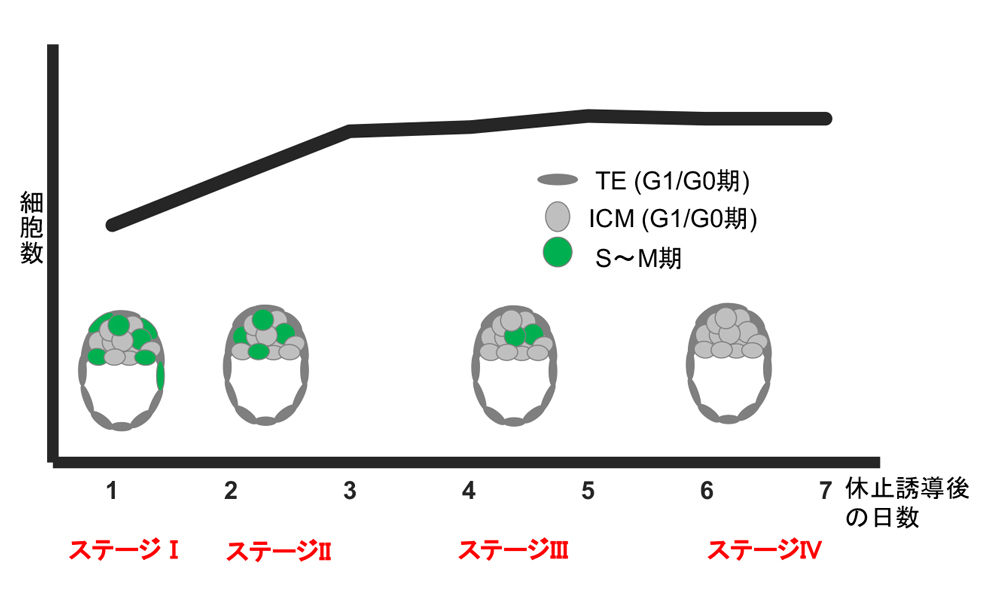 図1 発生休止は多段階的に進行する
図1 発生休止は多段階的に進行する
細胞数と細胞周期の変化のまとめ。発生休止誘導後しばらくは、緩やかに胚の中の細胞数は増加するが、やがて増加がみられなくなる。発生休止誘導直後から、非胚側の栄養外胚葉(TE)はG1/G0期に入る(ステージI)。胚側のTE細胞もG1/G0期に入り(ステージII),ステージIIIでは細胞数の増加が少なくなり多くの細胞がG0期に入る。やがて、細胞数の増加も見られなくなり全ての細胞がG0期に入る。
発生休止からの発生の再開に関して、胚盤胞の中で領域や細胞の分化状態によって違いがあるかについて調べる為に、発生休止中のR26-Fucci2マウス胚を培養し、Fucci2マーカーの発現を観察した。それぞれの休止段階から回収した胚を24時間培養して観察を行うと、全ての胚において胚側の領域で緑色の蛍光が見られた。休止後7日目の胚の培養では他の胚に比べて緑色へと変化した細胞数は少なかった。詳細な変化を確認するために、休止後3日目、7日目の胚を培養し、タイムラプス観察を行った。核の蛍光が赤色から緑色へと変化することが観察され、培養開始から核の色の変化に要した時間は休止誘導後7日目の胚の方が長い傾向がみられた。これらの結果から、発生休止胚が発生を再開する際には、胚側から再開し非胚側へと広がること、発生休止期間が長くなると発生再開に要する時間も長くなることが示唆された。
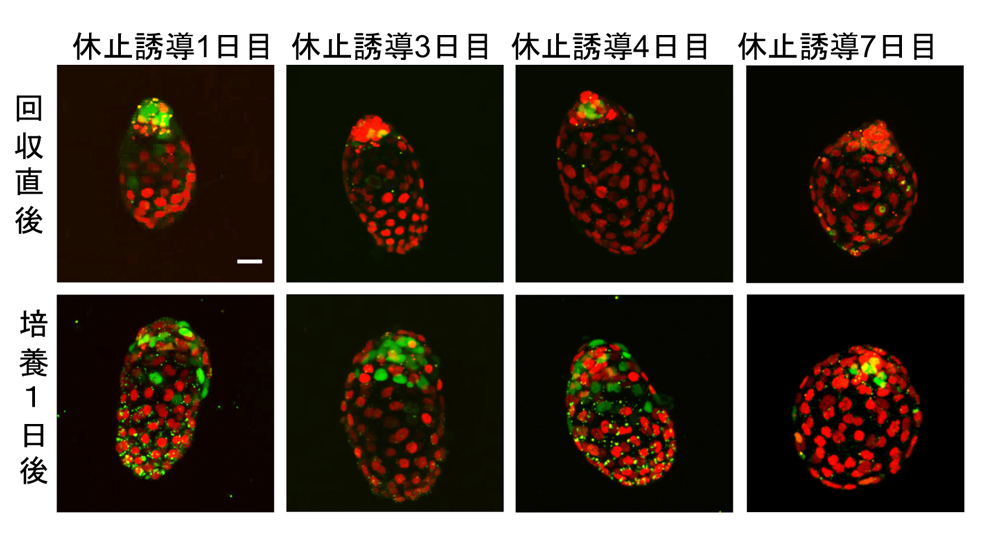 図2 休止胚での細胞周期の変化
図2 休止胚での細胞周期の変化
細胞周期を可視化することが可能な、R26pFucci2マウス胚を用いた観察。発生休止誘導後1,3,4,7日目の胚を回収して直後(上段)と、培養によって発生休止から退出して1日後の像。SからM期にある細胞の核は緑色、G1,G0期にある核は赤色の蛍光タンパク質で可視化される。休止誘導後1日目には、胚側領域だけでなく、非胚側領域の一部のTE細胞においても緑色が見られる。休止期間が長くなるとほぼ全ての細胞が赤色でのみ標識されることから、細胞周期が停止していることが示唆される。培養によって、発生休止から退出する際には、胚側領域から細胞周期が回り始め、やがて胚全体へと広がる。休止期間が長い胚は、この細胞周期の回復にも時間を要する。
以上の結果から、マウス胚の発生休止は段階的に起きることが示された。また、発生休止誘導後には、非胚側から細胞周期が緩やかに止まり、やがて胚全体へと広がり、この間それぞれの細胞の分化形質は維持されたままである。発生休止胚が発生を再開する際には逆に胚側から細胞周期が再開することも示された。上記の観察から、発生休止シグナルを胚が受ける際には、着床の際に直接子宮上皮と接触する非胚領域から胚全体へと休止状態が伝搬することが推測された。
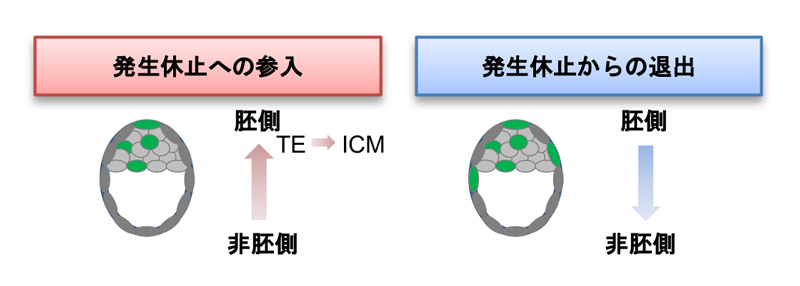 図3 胚の中の領域、分化形質による発生休止参入、退出のタイミングが異なる。
図3 胚の中の領域、分化形質による発生休止参入、退出のタイミングが異なる。
発生休止誘導後、非胚側のTE細胞から細胞周期が停止し、胚側のTE細胞、胚側のICM細胞の順で細胞周期は停止する。発生休止から解放された際には、胚側領域から細胞周期が回復しやがて胚全体で細胞周期が回り始める。
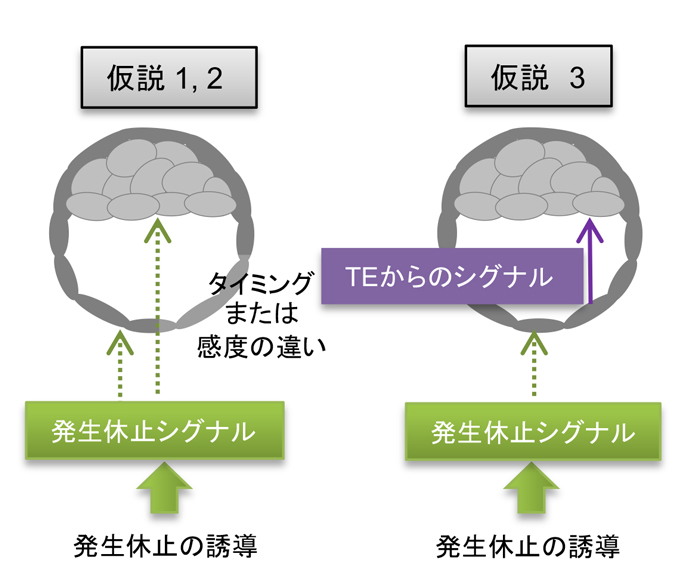 図4 発生休止シグナルの受容・反応に関する仮説
図4 発生休止シグナルの受容・反応に関する仮説
子宮からの発生休止シグナル(着床を受け入れないという状況)に対して、胚の領域によって細胞間で反応するタイミングが異なる。受容のタイミングや感度が領域によって異なる可能性(仮説1,2)、一旦非胚側のTEが受容したシグナルを別のシグナルとして放出し、それを胚領域で受容するという2段階のシグナルを介した可能性(仮説3)が考えられる。
【今後の展望】
本研究によって、発生休止への侵入と発生の再開が胚の領域間、分化形質間によって異なることが明らかになった。着床直後から胚の細胞は活発に細胞分裂を繰り返して、胚の大きさを増大する。発生休止においては、このような活発な活動が見られず、細胞が分化形質を維持した状態で緩やかに細胞周期を停止することが分かった。発生休止のそれぞれの段階で、胚の領域毎に遺伝子発現や細胞内の状態を解析すれば、細胞が休止するメカニズムの理解へとつながると期待している。成体での細胞の多くは分化形質を維持したまま、細胞分裂の起こらない状態にあり、傷などによって細胞周期が再開する。発生休止と成体での恒常性の維持に共通するメカニズムがあるかなど、興味深い課題であり今後の解明が期待される。
【掲載誌情報】
雑誌名: Biology of Reproduction
論文タイトル: Distinct dormancy progression depending on embryonic regions during mouse embryonic diapause
著者: Chizuru Kamemizu and Toshihiko Fujimori
DOI:
https://doi.org/10.1093/biolre/ioz017
【研究サポート】
本研究は文部科学省科学研究費助成事業およびJST戦略的創造研究推進事業の支援を受けて行われました。
【問い合わせ先】
基礎生物学研究所・初期発生研究部門
教授 藤森 俊彦(フジモリ トシヒコ)
〒444-8787 愛知県岡崎市明大寺町字東山5−1
TEL: 0564-59-5860
E-mail: fujimori@nibb.ac.jp
【報道担当】
基礎生物学研究所 広報室
TEL: 0564-55-7628
FAX: 0564-55-7597
E-mail: press@nibb.ac.jp




