基礎生物学研究所



2014.06.11
ミドリゾウリムシは、ゾウリムシと近縁の原生動物で、その細胞内にクロレラを共生することが知られています。ミドリゾウリムシはクロレラに二酸化炭素や窒素分を与え、クロレラは光合成を行い、光合成で得られた酸素や糖をミドリゾウリムシに与え、互いにメリットをもたらす「相利共生」の関係にあります。ミドリゾウリムシとクロレラは、真核細胞同士の細胞内共生(二次共生)の成立機構の解明に有用な研究材料として有望視されていますが、遺伝子に関する情報がほとんどありませんでした。今回、島根大学の児玉有紀准教授、山口大学の鈴木治夫准教授、藤島政博教授らは、基礎生物学研究所の重信秀治特任准教授と共同で、ミドリゾウリムシの大規模な遺伝子カタログを構築するとともに、クロレラとの共生の有無によって、ミドリゾウリムシの遺伝子発現がどのように変化するかを初めて明らかにしました。今回の研究成果を基盤に、ミドリゾウリムシが、共生研究のモデル系としてさらに活躍することが期待されます。本成果は、3月10日に科学雑誌「BMC Genomics」に掲載され、アクセス数が多い注目論文としてHighly accessed articleの認定を受けました。
【研究の背景】
ミドリゾウリムシ(Paramecium bursaria)の細胞内には約700個のクロレラ(Chlorella variabilis)が共生しています。光合成を行うことのできるクロレラは宿主のミドリゾウリムシに酸素やマルトースなどの光合成産物を与え、宿主はクロレラにCO2やNH3などを与えています。このため、ガス交換不能の密閉された容器にミドリゾウリムシを入れても、適度な光さえあれば、ミドリゾウリムシは酸欠や飢餓に耐え、長期間の生存が可能になります。このように、ミドリゾウリムシとクロレラの関係は「相利共生」と呼ばれるお互いにメリットをもたらす関係です。両者は単独で増殖することも可能であり、単独培養した両者を混合するとミドリゾウリムシの食胞を経由してクロレラとの再共生を容易に誘導できます(図1)。さらに、培養や、顕微鏡観察、実験操作のしやすさから、真核細胞同士の細胞内共生の成立機構を解明するための共生研究の新しいモデルとして有望視されてきました。実際、著者らによって、ミドリゾウリムシとクロレラの二次共生の経時的変化の追跡が可能になり(Kodama and Fujishima, Protoplasma 225, 191–203, 2005)、共生成立に必須な4つのステップが明らかになりました(Kodama and Fujishima, Int. Rev. Cell Mol. Biol., 279, 33–77, 2010)。このように、ミドリゾウリムシとクロレラの共生の現象面での理解は大きく進展した一方で、これまでミドリゾウリムシの遺伝子情報はほとんどなく、遺伝子や分子レベルでの研究は困難な状況にありました。
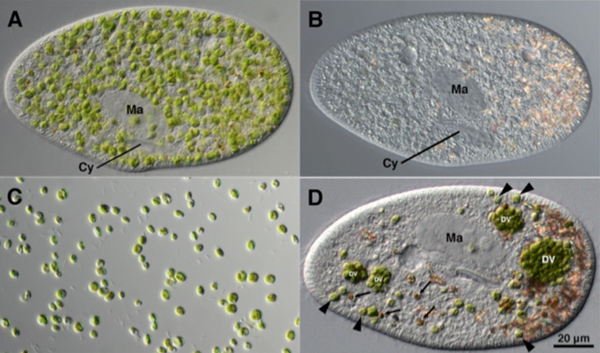
図1 A: クロレラが共生しているミドリゾウリムシ。B: Aを暗闇で培養することでクロレラを除去したミドリゾウリムシ。C: ミドリゾウリムシから単離した共生クロレラ。D: BとCを混合してから3時間後の細胞の様子。ミドリゾウリムシの食胞から脱出して再共生に成功したクロレラ(矢尻)が観察できる。Ma:大核、 Cy:細胞口、 DV:食胞 (Kodama and Fujishima, Protist, 163, 658–670, 2012から許可を得て転載。) なお、繊毛虫の仲間であるミドリゾウリムシはユーグレナ目に属するミドリムシとは異なる生物です。
【本研究の成果】
これまでミドリゾウリムシの遺伝子に関する情報はほとんどなかったので、研究グループは、まず、ミドリゾウリムシの網羅的な遺伝子カタログを作成しました。ミドリゾウリムシからRNAを抽出し、次世代シーケンサーを利用したRNA-seqと呼ばれる技術で40Gbp(400億塩基対)以上もの転写産物の塩基配列情報を取得し、大型計算機を用いて、10,557個のミドリゾウリムシ遺伝子を同定しました。
次に、クロレラを共生しているミドリゾウリムシとクロレラを除去したミドリゾウリムシからそれぞれRNAを抽出し、クロレラとの共生の有無により宿主であるミドリゾウリムシの遺伝子発現がどのように異なるのかを、上述した遺伝子セットをもとに調べました。発現が変化するミドリゾウリムシの遺伝子数は6,698個でした(図2)。共生によって発現量が変化する遺伝子群には、ストレスタンパク質遺伝子や抗酸化作用をもつグルタチオン-S-トランスフェラーゼ遺伝子などが含まれていました。これらの遺伝子が、共生の成立と維持にどのように関与しているのか、今後の研究が待たれます。
二次共生は細胞の進化の原動力となった生命現象で、地球上のいたるところで繰り返し起こっており、生命進化の重要な要因のひとつとして考えられています。今回の研究成果は、ミドリゾウリムシに二次共生研究のモデルとしての確固たる地位を与えただけでなく、多様な共生の成立機構を遺伝子レベルで理解する基盤となるでしょう。
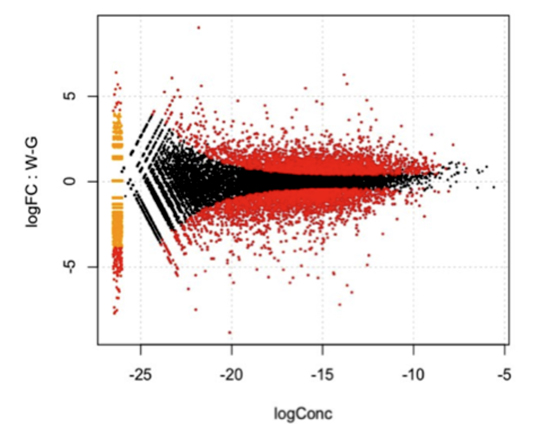
図2 クロレラ共生・非共生時の宿主ミドリゾウリムシの遺伝子発現変化。MAプロットと呼ばれるグラフ。ミドリゾウリムシのひとつひとつの遺伝子が点でプロットされている。X軸は遺伝子発現強度を示し、右に行くほど発現量が高いことを意味する。Y軸は発現の変動の倍率を示し、0は変動がないことを意味し、0から上下に乖離するほど変動が大きいことを意味する。赤のプロットは統計的に有意に発現の変動が認められた遺伝子。
注1)
RNA-seq: 次世代DNAシーケンサーを用いて、発現している遺伝子の転写産物を網羅的に解析する手法。次世代DNAシーケンサーは、高速並列シーケンスにより塩基配列情報をハイスループットに解読することができる装置。基礎生物学研究所は共同利用研究の一環として「次世代DNAシーケンサー共同利用実験」を実施しており、この論文の共著者の重信特任准教授はその責任者を務めている。
【論文情報】
BMC Genomics 2014年3月10日公開
論文タイトル:
著者:Y. Kodama, H. Suzuki, H. Dohra, M. Sugii, T. Kitazume, K. Yamaguchi, S. Shigenobu, and M. Fujishima (YK and HS contributed equally.)
【研究グループ】
この論文の第一著者は、児玉有紀准教授(島根大学生物資源科学部)と鈴木治夫准教授(山口大学大学院理工学研究科)で、文部科学省の特別経費「細胞内共生成立の分子機構の解明と新機能細胞の創成」(代表 山口大学大学院理工学研究科教授 藤島政博)の研究費の支援のもとで行われました。また、本研究は、基礎生物学研究所次世代DNAシーケンサー共同利用実験(担当:重信准教授)の一環として実施されました。
【本件に関するお問い合わせ】
島根大学 生物資源科学部 生物科学科
准教授 児玉 有紀(コダマ ユウキ)
〒690-8504 島根県松江市西川津町1060
TEL/FAX: 0852-32-6438 E-mail: kodama@life.shimane-u.ac.jp
山口大学 大学院理工学研究科
准教授(特命)鈴木 治夫 (スズキ ハルオ)
〒753-8512 山口市吉田1677-1
TEL: 083-933-5794 E-mail: haruo@yamaguchi-u.ac.jp
基礎生物学研究所 生物機能解析センター
特任准教授 重信 秀治(シゲノブ シュウジ)
〒444-8585 愛知県岡崎市明大寺町字西郷中38
Tel: 0564-55-7670 Fax: 0564-55-7669 E-mail: shige@nibb.ac.jp
山口大学 大学院理工学研究科 環境共生系学域
教授 藤島 政博 (フジシマ マサヒロ)
〒753-8512 山口市吉田1677-1
TEL/FAX: 083-933-5712 E-mail: fujishim@yamaguchi-u.ac.jp
【報道担当】
基礎生物学研究所 広報室
TEL: 0564-55-7628
FAX: 0564-55-7597
E-mail: press@nibb.ac.jp

