大山 隆
(早稲田大学教育・総合科学学術院・教授)
研究分担者
三谷 匡
(近畿大学・先端技術総合研究所・准教授)
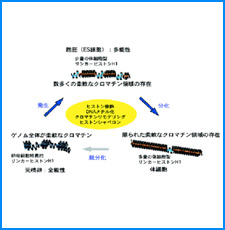
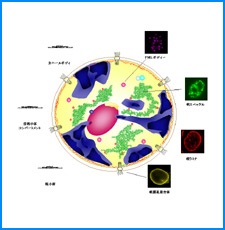
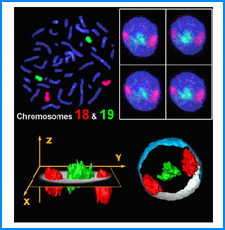
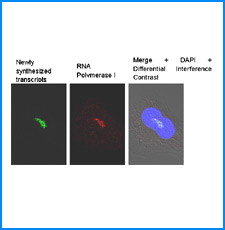
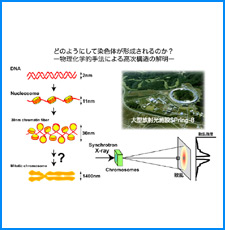
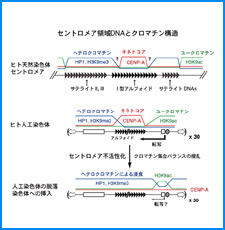
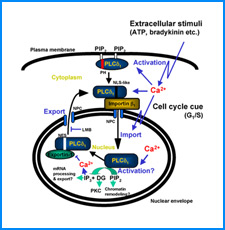
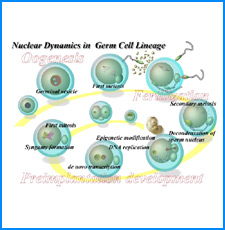
ゲノムを細胞核に機能的に折り畳むための遺伝情報はまだ解明されていない。しかし、これまでの研究でこの問題に関係すると思われる興味深い知見をいくつか得た。そこで、それらを手がかりとして、今後2年間で以下の研究を実施する。
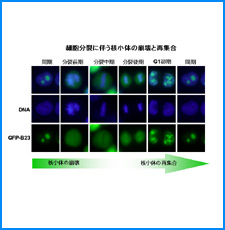
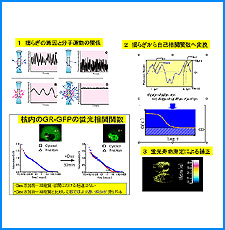
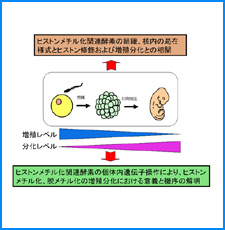
(i) 培養細胞を用いたこれまでの研究で、負の超らせんを擬態したベントDNA構造はクロマチン制御能と遺伝子発現制御能を有していることが明らかになった。そこで今回、ベントDNAのこのような機能が細胞分化や個体発生の過程で維持されるかどうかを調べる。
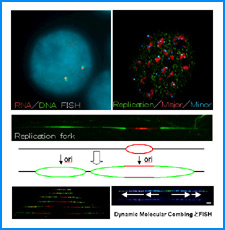
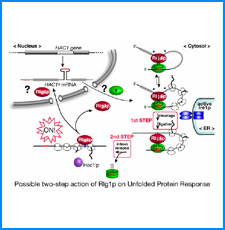
(ii) 我々は最近、DNAの機械的特性にも遺伝情報が印されていることを発見した。そこで今回、ヒトゲノムDNAの機械的特性地図を作成し、遺伝子の機能的折り畳み機構ならびに染色体の構築機構について考察する。
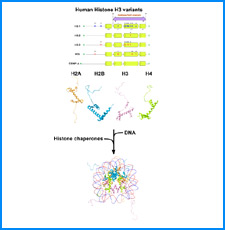
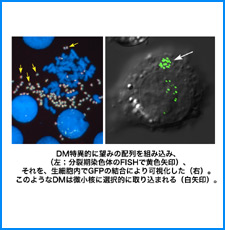
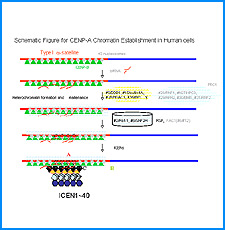
(iii) 我々は最近、DNAにも自己集合する性質があることを明らかにした。そこで今回、同じ配列のDNAをもつヌクレオソームに自己集合能があるかどうかを調べる。

|