広島大学 林 利憲(新規モデル生物共同利用研究課題代表者)
鳥取大学 竹内 隆(統合イメージング共同利用研究課題代表者)
基礎生物学研究所 亀井 保博(所内対応者)
両生類のイモリは、これまでに知られている脊椎動物の中で最も強い再生能力をもつことから、古くから器官再生の研究に用いられてきた。再生能力に加えて、イモリはヒトの約10倍の巨大ゲノムをもつ、がん化に対する耐性を有する、フェロモンを介した交尾行動をするなど、生物学的に興味深い性質を併せ持つ。しかし、これまで広く用いられてきたイモリの種は研究室内での繁殖が困難であり、イモリをモデル生物として使用できるような研究システムは整備されてこなかった。そこで我々は、イモリの新規のモデル生物として確立することを目指して、大量繁殖が容易なイベリアトゲイモリを導入した。さらに、基礎生物学研究所の新規モデル生物共同利用研究等による超階層生物学センターの支援を受けて、遺伝情報の整備(iNEWT・Blast検索サイト:
https://biaswiki.nibb.ac.jp/blast/blast.html)、ゲノム編集技術の確立、イメージング法の確立などに成功してきた。その結果、イベリアトゲイモリのユーザーや、本種を用いた科学論文の数は増加している。そこで、最近公表された本共同利用研究に関連する成果の一部(3報)を紹介する。

図1 イベリアトゲイモリの野生型(左)と色素変異体(右)。1回の産卵で数百個の受精卵を得ることができる。通年の繁殖が可能であり、メスは2、3週間ごとに産卵を繰り返す。
(1)イモリ膵臓の形態とその形成に関する研究
諸角 涼介、岡本 和子、林 利憲(広島大学両生類研究センター)
論文
Morozumi, R., Okamoto, K., Enomoto, E., Tsukamoto, Y., Kyakuno, M., Suzuki, N., Tazawa, I., Furuno, N., Ogino, H., Kamei, Y., Matsunami, M., Shigenobu, S., Suzuki, K., Uemasu, H., Namba, N., and Hayashi, T. (2025) Urodele amphibian newt bridges the missing link in evo-devo of the pancreas.
Dev. Dyn. doi:
10.1002/dvdy.763.
研究の概要
脊椎動物において膵臓は、消化と血糖調節の2つの機能をもつ重要な器官である。膵臓の疾患や異常は生存にとって重大な影響を及ぼすため、疾患の治療法や再生医療に対する関心も高い。特に、イモリの膵臓が再生する機構を解明できれば、ヒトの膵臓再生に応用する道が開かれる。しかし、膵臓の構造や機能は動物ごとに異なる点も多いことから、生物学的な視点だけでなく、イモリを再生医療や疾患のモデル動物として使用するためにも、膵臓に関する基本情報を得る必要がある。そこで本研究ではイベリアトゲイモリの膵臓の構造と発生過程の解析を行なった。その結果、イモリの膵臓は哺乳類と同様に一つの独立した器官として形成されることを示した。腸管上に膵臓様の組織を持つこと、この組織が有尾両生類固有のものであることを発見した。また、脊椎動物の膵臓形成の鍵となる遺伝子である
Pdx1 遺伝子に加えて、これまで一部の魚類にしか見つかっていない
Pdx2 遺伝子をイモリが有することを発見し、これらの機能を解析した。本研究は脊椎動物の膵臓が原始的な消化機能から血糖値調節担う複合的な器官へと進化する過程を理解する上で重要な成果となった。
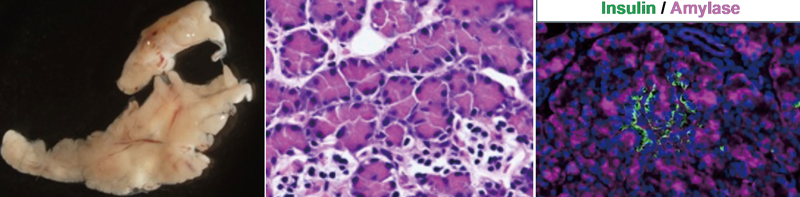
図2 イベリアトゲイモリ膵臓の全体像(左)と、ランゲルハンス島の組織像(中、右)。ランヘルハンス島では、哺乳類と同様にインスリンを合成する細胞が見られる(右)。
(2)ゲノム編集によるCdk1遺伝子の破壊がイモリ個体の細胞周期進行に与える影響の研究
中尾 勇太、岡本 和子、林 利憲(広島大学両生類研究センター)
論文
Nakao, Y., Okamoto, K., Tazawa, I., Nishijima, T., Furuno, N., Sakuma, T., Yamamoto, T., Takeuchi, T., Hayashi, T. (2024) Effect of
Cdk1 gene disruption on cell cycle progression in newt cells.
Dev. Growth & Differ. doi:
10.1111/dgd.12958.
研究の概要
細胞周期はCyclinとCyclin dependent kinase (CDK)によって制御される。脊椎動物では
Cdk1 のみが細胞増殖を進行させるために必須、すなわち他の
Cdk によって代替されない遺伝子であると考えられてきたもの、殆どの研究が哺乳類の細胞を用いて行われてきたため、Cdk1の機能が脊椎動物間でどの程度共通であるかはわかっていない。そこで本研究ではイベリアトゲイモリを用いて、
Cdk1遺伝子の破壊が細胞に与える影響を解析した。その結果、CRISPR/Cas9を介したゲノム編集により得た
Cdk1 破壊個体では、中枢神経系における細胞周期の早期停止が見られた。一方、肝臓と胃では細胞分裂期(M期)のスキップとDNAの再複製が起こったことが示された。本研究は、細胞増殖における
Cdk1 遺伝子の破壊の影響が、器官や細胞の種類ごとに異なることを明らかにした。
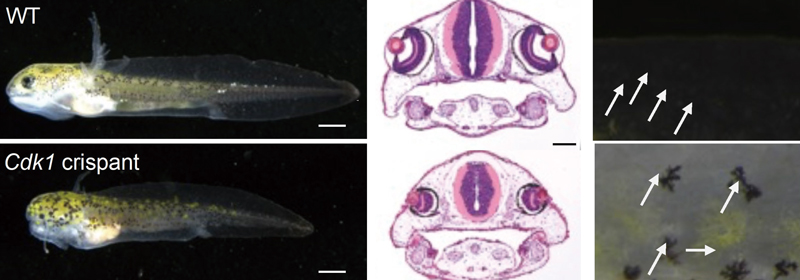
図3 イベリアトゲイモリの野生型(上段)と
Cdk1破壊(下段)幼生。幼生の外観(左)、頭部の組織像(中)、および体表の黒色色素細胞(右)。白矢印が示すように、
Cdk1を破壊した幼生では色素細胞が大型化している。
(3)イモリ後肢芽の伸長制御を行うHox13 の新たな機能
戸澤 紗代、竹内 隆 (鳥取大学医学部)
論文
Tozawa, S., Matsubara, H., Minamitani, F., Kamei, Y., Saida, M., Asao M., Suzuki, K. T., Matsunami, M., Shigenobu, S., Hayashi, T., Abe, G., and Takeuchi, T. (2025) Novel function of Hox13 in regulating outgrowth of the newt hindlimb bud through interaction with Fgf10 and Tbx4.
Dev. Growth & Differ. 67, 10-22. doi:
10.1111/dgd.12952. Epub 2024 Dec 26.
はじめに
Hox13遺伝子は前肢と後肢の両方において指の形成に必須であることがマウス変異体での研究から明らかになっていた。また、イモリにおいて発生および再生の両方においてHox13が指の形成に必要であることが私たちによって示された (Takeuchi et al, Development, 2022)。それに加えて、CRISPR/Cas9によるゲノム編集で作製されたHoxa13/c13/d13三重変異crispant(CRISPR/Cas9によって遺伝子破壊された個体)イモリの一部では、後肢の構造全体が欠損する表現型(以降、後肢欠損)が見つかった。この表現型は四肢において指の形成にのみ関わるとされたHox13遺伝子が後肢全体の形成に関わるという新機能ももつことを示している。そこでHox13遺伝子がどのように後肢全体の形成を行うのかを解析した。
結果
Hoxa13/c13/d13三重crispantイモリの一部で後肢全体が欠損する表現型(以降、後肢欠損)は、両肢もしくは片肢で認められた(図A)。イモリHox13遺伝子はa13, b13, c13,d13 の4つのパラログがあるが、後肢欠損は少なくともHoxa13, d13の二重変異が必要であった(図B)。また、生殖系列変異体(F2)においても、同様の結果を得た。このことからオフターゲット効果をほぼ否定できた。この生殖系列変異体でも片肢欠損が認められた。このことから後肢欠損の有り、無しは同一の遺伝子型かつ同一の遺伝的背景をもつ個体内でも生じる表現型であることが示された。さらに、基生研共同研究によりマイクロCTでの観察を行ったところ、欠損した肢においては大腿骨から先端の構造を全て失っていた(図C)。次に後肢の発生過程を観察したところ、Hox13 crispantイモリでは後肢芽は存在するが、伸長せず、退縮することがわかった。Whole-mount in situ hybridization(WISH)の結果から、欠損が予想される肢芽において、後肢伸長に関与する遺伝子Fgf10やTbx4の発現低下が認められた。これらFgf10、Tbx4それぞれのcrispantイモリを作製して観察した結果、その一部の個体でHox13変異イモリと同様の後肢の全欠損が認められた。最後に、Hox13/Fgf10, Hox13/Tbx4 多重crispantイモリを作製したところ、後肢欠損の割合がそれぞれの単独crispantと比較して統計的に有意に上昇した。このことから、Hox13とFgf10の間、また、Hox13とTbx4の間には遺伝学的相互作用があることが判明した。
考察
Hox13の変異による後肢欠損は、四肢動物においてイモリで初めて報告された表現型である。本研究の結果から、イモリHox13遺伝子が後肢の肢芽遠位側においてFgf10やTbx4の発現を亢進することで、肢芽の伸長を促すと考えられた。Hox13 変異体イモリ一部のみで後肢欠損が起こるのは、Fgf10やTbx4を制御する遺伝子がHox13以外にもあり、その機能が後肢ごとで変動しているためと考えた。進化過程で後肢を欠損した動物は多く知られており、本研究で明らかにされたHox13,Fgf10,Tbx4カスケードによる後肢形成機構との関係性に今後、興味が持たれる。
結論
Fgf10やTbx4との相互作用を通じてイモリの後肢芽の伸長を制御するというHox13 の新らたな機能が明らかにされた。
研究支援
本研究は基礎生物学研究所の共同利用研究(統合イメージング:マイクロCT解析:24NIBB525, 23NIBB503, 22NIBB504;新規モデル生物共同利用研究:イモリゲノム解析: 23NIBB201, 22NIBB201)の支援を受けて実施した。
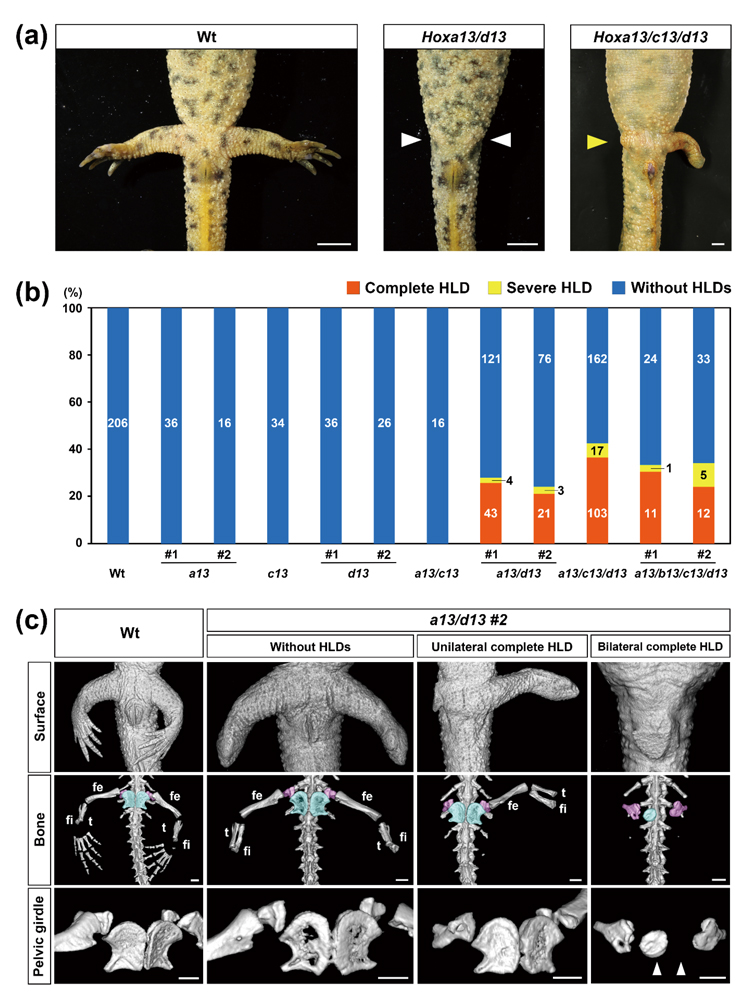
図4
(A)左と中央は、それぞれ受精後約5ヶ月の野生型イモリ(Wt)と後肢欠損を持つHoxa13/d13 クリスパントの腹側画像。右は受精後約1年における後肢欠損を有する同クリスパントの腹側画像。スケールバーは5mm。(B) 各遺伝子型における後肢欠損のないイモリ (without HLD)とあるイモリ(with HLD)の数の割合。それぞれの表現型を持つイモリの個体数を棒グラフ上で示した。異なるgRNAを用いて2つのクリスパント群(#1および#2)を作製した。後肢欠損のイモリの数には片側欠損と両側欠損の動物を含む。(C)WtイモリとHoxa13/d13クリスパントグループ個体#2(a13/d13 #2)のイモリの後肢と骨盤帯のマイクロCT画像(腹側)。中段と下段はそれぞれ、右斜め上から見た後肢と骨盤帯周辺の骨全体を表している。中段の水色とピンク色で示した部位は、それぞれ坐骨と腸骨を示す。feは大腿骨、fiは腓骨、tは脛骨。スケールバー、1mm。




