
サガリバナ科 Lecythidaceaeは約30属からなり、熱帯域に広く分布する(POWO 2026)。多数の雄ずいを形成するという共通の特徴をもち、雄ずい群の一部が仮雄ずい化する属や、立体的に多様な形態へと変化した属を含む。
Lecythidaceae comprises approximately 30 genera and is widely distributed in the tropics (POWO 2026). It is characterized by the production of numerous stamens and includes genera in which part of the androecium is modified into staminodes, as well as genera in which the androecium has evolved into highly three-dimensional and diverse forms.

(Napoleonaea leonensis: Photo by Carel Jongkind, https://www.inaturalist.org/photos/79271253, CC BY-NC
Scytopetalum pierreanum: Photo by Bart Wursten, https://www.inaturalist.org/photos/63908965, CC BY-NC
Foetidia mauritiana: Photo by marc_roussin, https://www.inaturalist.org/photos/223220378, CC BY-NC)

Phylogeny and divergence times of Lecythidaceae subfamilies
サガリバナ科は、5亜科を含む広義のサガリバナ科として分類される場合(APG IV 2016;Stevens 2001 onwards)と、ナポレオナエア亜科およびスキトペタルム亜科をそれぞれ独立の科とし、残りの3亜科からなる狭義のサガリバナ科として分類される場合がある(Huang et al. 2015;Mori et al. 2017)。本稿では広義のサガリバナ科を採用する。系統樹の樹形および分岐年代は Rose et al. (2018) に基づく。
Lecythidaceae has been classified either in a broad sense, Lecythidaceae sensu lato including five subfamilies (APG IV 2016; Stevens 2001 onwards), or in a narrow sense, in which Napoleonaeoideae and Scytopetaloideae are treated as separate families and the remaining three subfamilies constitute Lecythidaceae sensu stricto (Huang et al. 2015; Mori et al. 2017). In this study, Lecythidaceae is treated in the broad sense. The topology and divergence times of the above phylogeny follow Rose et al. (2018).
サガリバナ科のマクロ形態に基づく検索表
1. 仮雄蕊が花弁化する …… ナポレオナエア亜科Napoleonaeaceae
1. 仮雄蕊は花弁化しない…… 2
2. 子房は上位またはやや下位 ……スキトペタルム亜科 Scytopetaloideae
2. 子房は下位または半下位 …… 3
3. 花弁を欠く …… フォエティディア亜科Foetidioideae
3. 花弁をもつ …… 4
4. 雄ずい群の内側に環状蜜腺あり;旧熱帯 ……サガリバナ亜科Barringtonioideae
4. 雄ずい群の内側に環状蜜腺なし;新熱帯 …… パラダイスットノキ亜科Lecythidoideae
Dichotomous key to the Lecythidaceae based on macromorphology
1. Staminodes petaloid … Napoleonaeoideae (Napoleonaeaceae)
1. Staminodes not petaloid … 2
2. Ovary superior or semi-inferior … Asteranthoideae
2. Ovary inferior or half-inferior … 3
3. Petals absent … Foetidioideae
3. Petals present … 4
4. Annular intrastaminal nectary present; distribution in the Old World … Barringtonioideae
4. Annular intrastaminal nectary absent; distribution in the New World … Lecythidoideae

Napoleonaea and Crateranthus in Napoleonaeoideae
Napoleonaea beninensis: Photo by Scott Zona, https://www.inaturalist.org/photos/91855160, CC BY-NC
ナポレオナエア亜科はナポレオナエア属Napoleonaeaとクラテランテゥス属Crateranthusの2属を含む(Mori et al. 2017)。多数の雄ずいを形成し、その一部が花弁状の仮雄ずいとなる点が共通の特徴がある。写真では、花弁化した仮雄ずいを赤色の矢印で示した(図1、2)。Crateranthus latestuiの線画(図2)の説明 (Lecomte 1920):1. 葉をつけた枝; 2. つぼみ; 3. 分離した萼片; 4. 萼片を除去して仮雄ずい性コロナ(紫色)を示した花; 5. 同、上方から見たもの; 6. 開花前の花の縦断面; 7. 開花後の花の縦断面; 8. 子房部における横断面; 9. 仮雄ずい性コロナの一部(縁の一部が持ち上がった状態); 10. 葯の裂開前および裂開後の雄ずい。
Napoleonaeoideae comprises two genera, Napoleonaea and Crateranthus (Mori et al. 2017). It is characterized by the production of numerous stamens, some of which are modified into petaloid staminodes. In the photographs, the petaloid staminodes are indicated by red arrows (Figs. 1, 2). Caption of the line drawing of Crateranthus latestui (Fig. 2) (Lecomte 1920): 1. Branch with leaves; 2. Bud; 3. Detached sepal; 4. Flower with sepals removed to show the staminodial corona (purple); 5. Same, viewed from above; 6. Longitudinal section of the flower before anthesis; 7. Longitudinal section of the flower after anthesis; 8. Transverse section of the ovary region; 9. Part of the staminodial corona (margin partly raised); 10. Stamens before and after anther dehiscence.

Distribution of the two genera of Napoleonaeoideae, Napoleonaea and Crateranthus
POWO 2026: Plants of the World Online © Board of Trustees of the Royal Botanic Gardens, Kew: http://www.plantsoftheworldonline.org/ (accessed 4 May 2026). CC BY 4.0.
2属ともアフリカ西部からアフリカ南部にかけて分布する(POWO 2026)。
Both genera are distributed from West Africa to Central Africa.

A flower of Napoleonaea beninensis
Napoleonaea beninensis: Photo by Scott Zona, https://commons.wikimedia.org/wiki/File:Napoleonaea_imperialis-2.jpg, CC BY 2.0.
ナポレオナエア属Napoleonaeaの花は、最も複雑な構造をもつ花の一つと考えられている(Frame and Durou 2001) (図5)。本属の特徴は、5裂した扁平な柱頭をもつことである(図6)。Napoleonaea beninensisの写真は記載の無いものは、名古屋市東山植物園の栽培株を2026年4月22日に撮影させていただきました。心より感謝申し上げます。
The flowers of Napoleonaea are considered to be among the most structurally complex (Frame and Durou 2001) (Fig. 5). A distinctive feature of the genus is the presence of a broad, flattened, five-lobed stigma (Fig. 6). Unless otherwise noted, photographs of Napoleonaea beninensis were taken from a cultivated plant at Nagoya Higashiyama Botanical Garden on April 22, 2026. I sincerely thank them for their excellent cultivation.

Floral organs of Napoleonaea beninensis
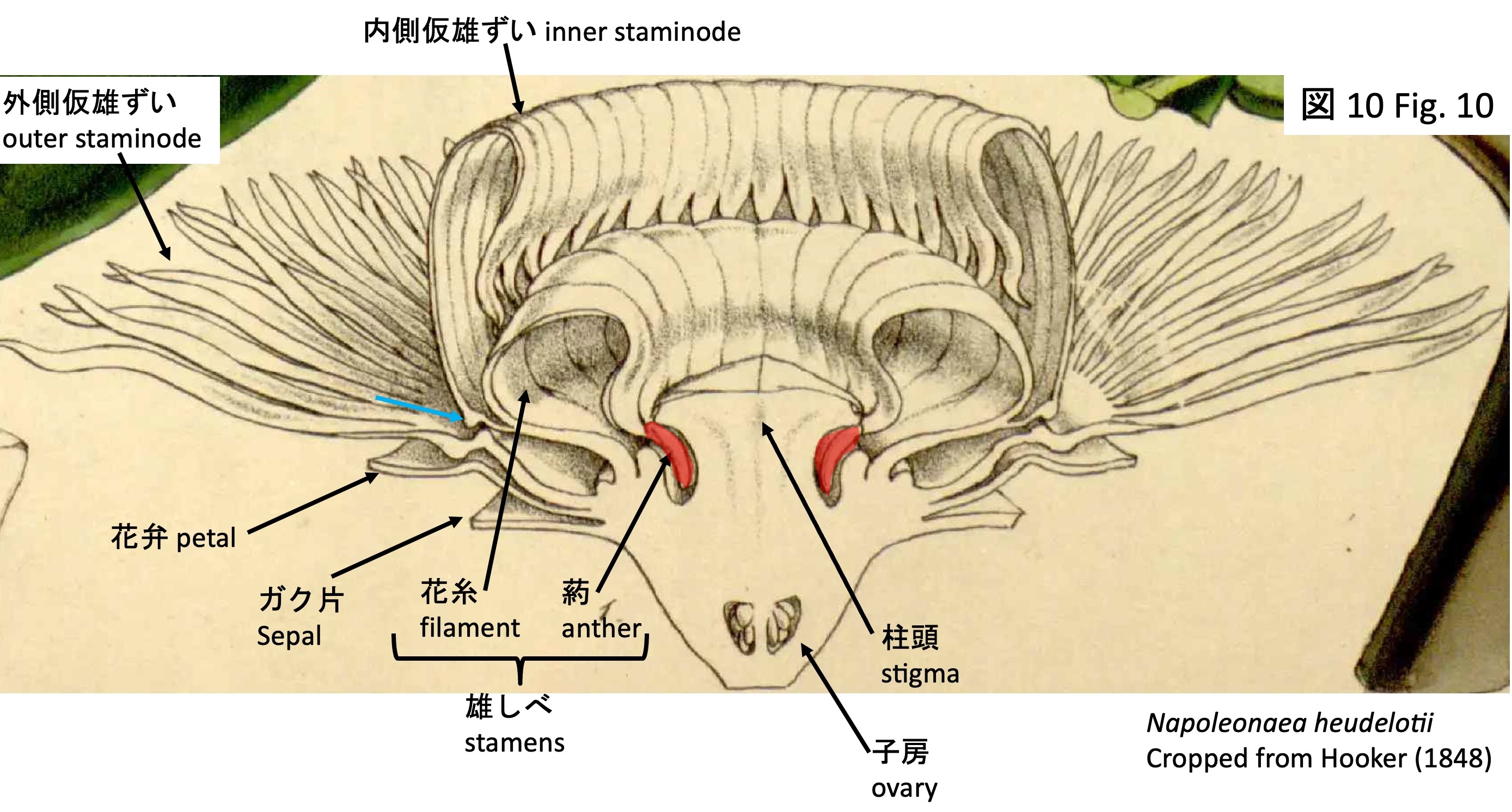
Longitudinal section of the flower of Napoleonaea heudelotii (identified as Napoleonaea imperialis in Hooker 1848, but later corrected to N. heudelotii by Prance and Jongkind [2015]).
Napoleonaea imperialisとN. beninensisの花の発生過程を調べた研究(Ronse de Craene 2011;原著でN. vogeliiとされているものは、Prance and Jongkind 2015によればN. beninensisに相当する)によると、花形成初期には数枚の小苞葉(図7)が形成され、その内側に生じる花原基から、まず5枚の萼片(図7)が輪状に形成される。次に、その内側に花弁(図8)が輪状に形成される。
花器官の原基形成順序は多様であり、典型的には萼片、花弁、雄ずい、雌ずいの順に形成されるが、雄ずいが花弁に先行する(萼片、雄ずい、花弁、雌ずい)場合も少なくない(Gifford and Foster 1989)。ところが、ナポレオナエア属では、花弁原基が伸長した後、雄ずいではなく花原基中央から雌ずい(図6)が形成される。その後、花弁と雌しべの間に雄ずい原基が輪状に伸長する。
雄ずい(図9)は、葯が発達する2本の稔性雄ずい(図9のF)、葯が発達しない不稔雄ずい2本(図9のS)とが交互に配置されて形成される。通常、稔性雄ずい、不稔雄ずいともに、扁平な5角形の柱頭の下側に潜り込むように形成されるが(図10のN. heudelotiiの花の断面図では、柱頭下に入り込む葯を赤色で示した)、本栽培株では柱頭の発達が不十分なのか、雄ずいが露出している(図9)。
その後、雄ずい原基の外側に内側仮雄ずい、続いてその外側に外側仮雄ずいの原基が伸長する。
クラテランテゥス属Crateranthusの花(図2)については、発生過程の報告はない。花弁は形成されず、花弁状の仮雄ずいが形成されると考えられている(Lecomte 1920, Prance and Jongkind 2015)。この構造が、ナポレオナエア属の花弁状仮雄ずいと相同であるならば、ナポレオナエア属と同様に、雌ずい原基の発達後に形成される可能性が高い。一方、もし雌ずい原基よりも早期に形成されるのであれば、花弁に相同な可能性も考えられる。
しかし、ナポレオナエア属の内側仮雄ずいに見られるような背側の突起(図2と図10の水色矢印)が存在することから、両者は相同な器官である可能性が高いと考えられる。この突起の機能については、両属ともに明らかになっていない。
Based on a study of floral development in Napoleonaea imperialis and N. beninensis (Ronse de Craene 2011; the material identified as N. vogelii in the original paper corresponds to N. beninensis according to Prance and Jongkind 2015), several bracteoles (Fig. 7) are first formed at an early stage of flower development. From the floral primordium arising inside these, five sepals (Fig. 7) are initiated in a whorl, followed by the whorled initiation of petals (Fig. 8).
The sequence of floral organ initiation is variable. Typically, organs arise in the order sepals, petals, stamens, and carpels, but in many cases stamens precede petals (sepals, stamens, petals, carpels). In Napoleonaea, however, after the petal primordia begin to elongate, the carpel primordia (Fig. 6) are initiated from the center of the floral primordium before the stamen primordia. Subsequently, stamen primordia develop in a whorl between the petal and carpel primordia.
The stamens (Fig. 9) are arranged as neighboring pairs of fertile stamens with developed anthers (F in Fig. 9) and sterile stamens lacking anthers (S in Fig. 9). Typically, both fertile and sterile stamens are positioned so as to lie beneath the flat, pentagonal stigma; in a sectional view of N. heudelotii (Fig. 10), the anthers inserted beneath the stigma are indicated in red. In the cultivated material examined here, however, the stigma is poorly developed, and the stamens are exposed (Fig. 9).
Subsequently, inner staminode primordia and then outer staminode primordia arise outside the stamen primordia.
No studies have reported the floral development of Crateranthus (Fig. 2). Petals are considered to be absent, with petaloid staminodes being formed instead (Lecomte 1920; Prance and Jongkind 2015). If these structures are homologous to the petaloid staminodes of Napoleonaea, they are likely to arise after the development of the carpel primordia, as in Napoleonaea. Conversely, if they are initiated earlier than the carpel primordia, they may be homologous to Napoleonaea petals.
However, the presence of dorsal projections (light blue arrows in Figs. 2 and 10), similar to those observed in the inner staminodes of Napoleonaea, suggests that these structures are most likely homologous. The function of these projections remains unknown in both genera.

Pollinators of Napoleonaea angolensis
Napoleonaea angolensis-1: Photo by Bart Wursten, https://www.inaturalist.org/photos/63897726, CC BY-NC
Napoleonaea angolensis-2: Photo by Anne-Hélène Paradis, https://www.inaturalist.org/photos/615702496, CC BY-NC
ナポレオナエア属の花が複雑化した理由として、送粉昆虫との共進化が考えられる。Napoleonaea angolensisについては訪花昆虫が調べられている(図11、12)(Frame and Durou 2001; 注:Frame and Durou [2001]ではN. vogeliiとして記述されたが、Prance and Jongkind (2015)でN. angolensisに訂正された。)。
蕾の段階では、萼片先端部の蜜腺から蜜が分泌されており(図13、14にN. beniensisの写真を示す)、これをアリ類が採取する。アリ類の誘引により若い花芽を食害昆虫から防御する効果が期待されるが、自生地では約1/3の蕾がイモムシに食害されており、その効果の程度は明らかではない。
花は19時頃から開くが、この時間帯には訪花昆虫は観察されていない。翌朝8時頃になると花はチョコレート様の香りを発し、それに伴って昆虫の訪花が始まる。アザミウマ類は、花香に含まれるアルデヒド類や白色に誘引されること、柱頭と雄ずいの間に形成される空間(花室:floral chamber)は隠れ場所として適していること、さらに実際に花室内で体に花粉を付着させた個体が観察されていることから、送粉昆虫である可能性が高いと考えられている。
しかし、花室内にアザミウマ類を定着させる花は、Calluna(Hagerup 1950)やPeixotoa(Del-Claro et al. 1997)で知られているものの、ナポレオナエア属ほど複雑ではない。ナポレオナエア属の花がなぜこれほど複雑な構造をもつのかは、十分には説明されていない。
自生地では、観察した26個の花のうち、子房が膨らんだものは1個のみであり、結実率は低い。このことも踏まえると、過去にはアザミウマ類以外の送粉昆虫が存在し、それに適応していた可能性も考えられている(Frame and Durou 2001)。
The remarkable complexity of flowers in Napoleonaea is thought to reflect coevolution with pollinating insects. Insect visitors have been studied in N. angolensis (Figs. 11, 12) (Frame and Durou 2001; note: the material treated as N. vogelii in Frame and Durou [2001] was reassigned to N. angolensis by Prance and Jongkind [2015]).
At the bud stage, nectar is secreted from nectaries at the adaxial distal positions of the sepals (photographs of N. beninensis are shown in Figs. 13, 14), and ants are attracted to collect it. Attraction of ants is expected to provide some protection of young floral buds against herbivores; however, in natural populations about one-third of the buds are damaged by caterpillars, and the effectiveness of ant attraction remains unclear.
Flowers open around 7 p.m., but no insect visitors have been observed at that time. Around 8 a.m. the following morning, the flowers begin to emit a chocolate-like scent, and insect visitation increases. Thrips are attracted to aldehydes in the floral scent and to the white color; the space between the stigma and stamens (the floral chamber) provides a suitable refuge; and individuals bearing pollen have been observed within this chamber. These observations suggest that thrips are likely pollinators.
However, although flowers that harbor thrips within a floral chamber are known in Calluna (Hagerup 1950) and Peixotoa (Del-Claro et al. 1997), they are far less complex than those of Napoleonaea (Frame and Durou 2001). The reason for the extreme complexity of Napoleonaea flowers remains unclear.
In natural populations, only one out of 26 observed flowers showed ovary enlargement, indicating a low fruit set. Taken together, these observations raise the possibility that Napoleonaea may have originally been adapted to pollinators other than thrips (Frame and Durou 2001).
引用文献 References
Angiosperm Phylogeny Group (2016). An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG IV. Bot. J. Linn. Soc. 181, 1–20.
Del-Claro, K., Marullo, R., and Mound, L.A. (1997). A new Brazilian species of Heterothrips (Insecta: Thysanoptera) co-existing with ants in the flowers of Peixotoa tomentosa (Malphigiaceae). J. Nat. Hist. 31, 1307–1312.
Frame, D., and Durou, S. (2001). Morphology and biology of Napoleonaea vogelii (Lecythidaceae) flowers in relation to the natural history of insect visitors. Biotropica 33, 458–471. https://doi.org/10.1111/J.1744-7429.2001.TB00199.X;PAGE:STRING:ARTICLE/CHAPTER.
Gifford, E.M., and Foster, A.S. (1989). Morphology and Evolution of Vascular Plants 3rd ed. (W. H. Freeman and Company).
Hagerup, O. (1950). Thrips pollination in Calluna. Danske Biol. Meddel. 18, 1–16.
Hooker, W.J. (1848). Curtis’s Botanical Magazine 74: Tab. 4387
Huang, Y.Y., Mori, S.A., and Kelly, L.M. (2015). Toward a phylogenetic-based generic classification of neotropical Lecythidaceae – I. Status of Bertholletia, Corythophora, Eschweilera and Lecythis. Phytotaxa 203, 85-121–185. https://doi.org/10.11646/PHYTOTAXA.203.2.1.
Kandidov, M.V., Bobrov, A.V., Kandidov, M.V., and Bobrov, A.V. (2026). The history of distribution of the family Lecythidaceae s. l. Geogr. Environ. Sustain. https://doi.org/10.24057/2071-9388-2026-4560.
Lecomte, M.H. 1920. Deux especes nouvelles de genre Crateranthus (Lecythidacees). Bull. Muséum National d’Histoire Naturelle 26: 68-71.
Mori, S.A., Kiernan, E.A., Smith, N.P., Kelly, L.M., Huang, Y.-Y., Prance, G.T., and Thiers, B. (2017). Observations on the phytogeography of the Lecythidaceae clade (brazil nut family). Phytoneuron 30, 1–85.
POWO 2026: Plants of the World Online © Board of Trustees of the Royal Botanic Gardens, Kew: http://www.plantsoftheworldonline.org/ (accessed 4 May 2026). CC BY 4.0.
Prance, G.T., and Jongkind, C.C.H. (2015). A revision of African Lecythidaceae. Kew Bull. 70, 1–68.
Ronse de Craene, L.P. (2011). Floral development of Napoleonaea (Lecythidaceae), a deceptively complex flower. In Flowers on the Tree of Life, L. Wanntorp and L. P. Ronse de Craene, eds. (Cambridge Univ. Press), pp. 279–295.
Rose, J.P., Kleist, T.J., Löfstrand, S.D., Drew, B.T., Schönenberger, J., and Sytsma, K.J. (2018). Phylogeny, historical biogeography, and diversification of angiosperm order Ericales suggest ancient Neotropical and East Asian connections. Mol. Phylogenet. Evol. 122, 59–79. https://doi.org/10.1016/J.YMPEV.2018.01.014.
Stevens, P. F. (2001 onwards). Angiosperm Phylogeny Website. Version 14. http://www.mobot.org/MOBOT/research/APweb/.