
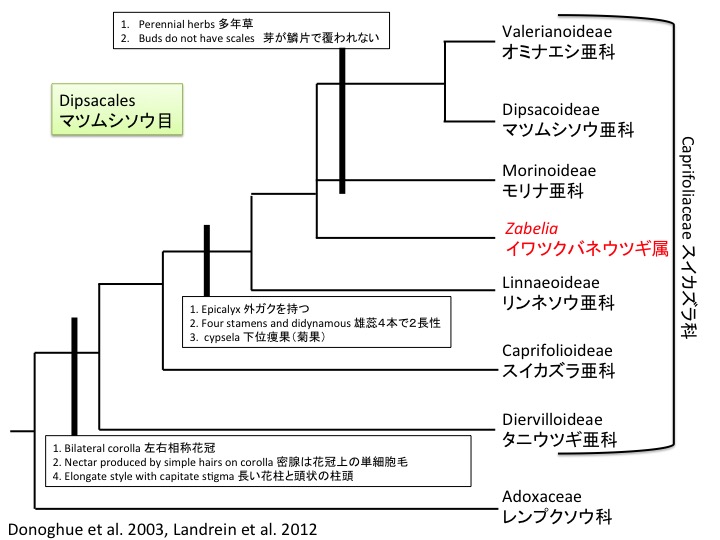


イワツクバネウツギ属の共有派生形質は葉柄基部が膨らみ腋芽を包むことである。どのような仕組みで膨らむのだろうか。また、何のために膨らむのだろうか。腋芽の保護のためなら、どうして他属でも同じように膨らまないのだろうか。
A synapomorphic character of Zabelia is the petiole bases swollen to enclose axillary buds. How and why the bases are swollen?

イワツクバネウツギ属の共有派生形質は葉柄基部が膨らみ腋芽を包むことである。どのような仕組みで膨らむのだろうか。また、何のために膨らむのだろうか。腋芽の保護のためなら、どうして他属でも同じように膨らまないのだろうか。
A synapomorphic character of Zabelia is the petiole bases swollen to enclose axillary buds. How and why the bases are swollen?
Actinotus フランネルフラワー属は19種がオーストラリアとニュージーランドに分布する(Henwood 2013)。
Actinotus is distributed in Australia and New Zealand (Henwood 2013).
Henwood, M.J. 2013. Actinotus repens Keighery ex Henwood (Apiaceae): A new species from south-west Western Australia. TELOPEA 15: 221-225.
柱下体は花筒に包まれずむき出しになっている。
Stylopodiua are not covered by hypanthium.


Apiopetalum属は2種からなりニューカレドニアの固有種で、大きくなると約6mほどの木になり(Oskolski and Lowry 2000)、セリ科よりもウコギ科に似ている。しかし、セリ科の共有派生形質である、かぎ爪状の花弁、2心皮性雌蕊、葉柄基部が茎全体を取り巻くという形質を持っており(Oskolski and Lowry 2000)、遺伝子配列比較からもセリ科であると推定されている (Nicolas and Plunkett 2014)。
Apiopetalum is composed of two species endemic in New Caledonia (Oskolski and Lowry 2000). Instead of the woody nature unusual in the Apiaceae and usual in the Araliaceae, some characters including the presence of clawed petals, a bicarpellate gynoecium, and leaf bases encircling a stem are shared with other species in the Apiaceae (Oskolski and Lowry 2000). Inclusion of Apiopetalum in the Apiaceae is supported by molecular phylogenetic analyses (Nicolas and Plunkett 2014).
Nicolas, A. N. and Plunkett, G. M. 2014. Diversification times and biogeographic patterns in Apiales. Bot. Review 80: 30-58.
Oskolski, A.A. and Lowry II, P.P. 2000. Wood Anatomy of Mackinlaya and Apiopetalum (Araliaceae) and Its Systematic Implications. Ann. Missouri Bot. Gard. 87: 171-182.


マッキンラヤ属は5種からなり(Philipson 1979)、オーストラリアからマレーシアに分布している(Nicolas and Plunkett 2014)。低木になる点はセリ科では木になるApiopetalumのように稀である。しかし、葉柄基部が茎を抱く点はセリ科の共有派生形質である。
Mackinlaya is composed of five species (Philipson 1979) and distribute from Australia to Malesia (Nicolas and Plunkett 2014). Although the shrubby morphology is unusual in the Apiaceae as the woody Apiopetalum, “leaf bases encircling a stem“ is a synapomorphic character of the Apiaceae.
Nicolas, A. N. and Plunkett, G. M. 2014. Diversification times and biogeographic patterns in Apiales. Bot. Review 80: 30-58.
Philipson, W.R. 1979. Araliaceae, part 1. Ed. C.G.G.J. van Steenis, Flora Malesiana, Ser. I, 9: 1-105. Martinus Nijhoff, Dr. W. Junk Publishers, The Hague, Boston, London.

さらに、舷部のある花弁、心皮性の子房、そして、分子系統解析の結果もセリ科の一員であることを示している。
In addition, clawed petals, a bicarpellate gynoecium, and molecular phylogeny inference support its inclusion in the Apiaceae.


Xanthosia属は約20種からなり、オーストラリアに分布している。Xanthosia rotundifoliaの総苞は花弁化している。
Xanthosia is composed of about 20 species and is distributed in Australia. Xanthosia rotundifolia forms petaloid bracts.


ツボクサ属は約100種が世界中に分布する。
茎が這って節から根を出すことや腋生の花序がチドメグサ属 Hydrocotyleとツボクサ属 Centellaで似ているが、遺伝子解析から、チドメグサ属はウコギ科、ツボクサ属はセリ科に属することがわかった。
ツボクサ属はセリ科の以下の2つの共有派生形質を共有している:1つは葉柄基部が茎を取り巻くことである。
Although the creeping stem, axillary inflorescences, and adventitious roots from nodes are similar between Hydrocotyle and Centrella, Hydrocotyle belongs in the Araliaceae and Centella in the Apiaceae.
Centella share the following two synapomorphic characters of the Apiaceae: (1) Leaf bases encircle a stem.

もう一つは花弁辺縁が内側に曲がり、かぎ爪状であることである。しかし、セリ科に広く見られる子房の油管は無い。
「蕾の中で著しく曲がる雄しべ」はセリ科とミオドカルパス科の共有派生形質であるが、ツボクサ属の雄しべは蕾の中でそれほど曲がらない。
(2) Petals are clawed at the base and inflexed at the tips. On the other hand, Centella do not have oil canals in the fruits, although it is synapomorphic character of the Apiaceae.
“Extensively inflexed stamens in a flower bud” is a synapomorphic character of the Apiaceae and Myodocarpaceae, but stamens of Centella are not so inflexed.



Nicolas, A.N. and Plunkett, G.M. 2009. The demise of subfamily Hydrocotyloideae (Apiaceae) and the re-alignment of its genera across the entire order Apiales. Molec. Phylogenet. Evol. 53: 134-151.



チドメグサ属と関連属はセリ科に分類されてきたが、塩基配列情報を用いた系統推定によってウコギ科に属する可能性が高いことがわかった (例えば Nicolas and Plunkett 2009)。
Hydrocotyle and its close relatives have been treated as a member of the Apiaceae but phylogenetic analyses using nucleotide sequences support the inclusion in the Araliaceae (e.g. Nicolas and Plunkett 2009).
Nicolas, A.N. and Plunkett, G.M. 2009. The demise of subfamily Hydrocotyloideae (Apiaceae) and the re-alignment of its genera axross the entire order Apiales. Molec. Phylogenet. Evol. 53: 134-151.

チドメグサ属はいくつかの形態形質をウコギ科と共有しているが、他の形態形質はセリ科と共有している。似ている方に+の印を付けた。両科の中間的な形態をしている場合は点線の上に+印を付けた。
Hydrocotyle shares some morphological characters with the Araliaceae but other ones with the Apiaceae. Morphological characters of Hydrocotyle are indicated as plus marks. When Hydrocotyle has intermediate morphology, plus mark is shown on the dotted line.

ヤツデの托葉は葉柄と癒合している。
Stipules of Fatsia japonica is connate to a petiole.

ウコギ科のほとんどは木本性だがセリ科のほとんどは草本性である。チドメグサ属は草本である。
The Araliaceae are mostly composed of woody species, while the Apiaceae are of herbaceous species. Hydrocotyle is herbaceous.

ウコギ科では、それぞれの花柱の基部に形成される柱下体が癒合するが、セリ科は癒合しない。チドメグサ属は癒合しておらずセリ科に似ている。
Stylopodia at the base of styles are connate in the Araliaceae, but are not connate in the Apiaceae. Stylopodia of Hydrocotyle are not connate as that of the Apiaceae.





加えて、チドメグサ属はセリ科のツボクサ属に似ている。チドメグサ属の形態形質を緑色の+印、ツボクサ属の形態形質を赤色の+印で示した。
In addition, Centella in the Apiaceae is similar to Hydrocotyle. Morphological characters of Centella and Hydrocotyle are shown as red and green plus marks, respectively.


柱下体は両種とも癒合しないが、花柱の位置する場所が異なる。チドメグサ属では花柱は柱下体の中央部あたりに位置するが、ツボクサでは心皮の境界部分に位置する。
Stylopodia are not connate in both species, but position of styles are different between them. Styles are positioned at the center of each stylopodium in Hydrocotyle, while they are positioned at the border of two carpels in Centella.

チドメグサ属の花弁はウコギ科、ツボクサ属の花弁はセリ科の多くが持つ特徴に合致する。一方、雄蕊形態はやや曲がり、ともにウコギ科とセリ科の中間的な形態である。
Petals of Hydrocotyle and Centella are similar to those in the Araliaceae and the Apiaceae, respectively. Stamens of both genera are slightly bend and intermediate between both families.

托葉や葉の基部形態は二属で異なっている。
Morphology at the leaf base and stipules is different between two genera.

さらに、チドメグサ属とツボクサ属は、ウコギ科とセリ科の両者の中で稀な次の3つの形態形質を共有している。1)茎が長く這う点、2)匍匐茎から不定根を形成する、3)花序を腋生する。これらのことから、チドメグサ属はツボクサ属に近縁、すなわち、セリ科に属すると考えられてきた。ただ、よく観察するとチドメグサ属の花序は腋生ではない。
Furthermore, Hydrocotyle and Centella share the following three special morphological characters that are not popular in both Araliaceae and Apiaceae: 1) the creeping stem, 2) adventitious roots from nodes, and 3) axillary inflorescences. Therefore, Hydrocotyle was classified into the Apiaceae. However, with careful observation, inflorescences of Hydrocotyle are not axillary.

セリ科のツボクサ属では、花序は葉腋に形成され、蔓状のシュートは単軸分枝である。
Inflorescences of Centella in the Apiaceae are axillary. The creeping shoot is monopodial.
 アマゾンチドメグサの花序は葉の腋ではなく、葉と対生の位置に形成される。このことは、このシュートは仮軸分枝で、花序が頂端で、蔓の先端部は葉腋にできた腋芽である可能性を示唆している。右の写真では主腋芽と葉の間に副芽が形成されている。
アマゾンチドメグサの花序は葉の腋ではなく、葉と対生の位置に形成される。このことは、このシュートは仮軸分枝で、花序が頂端で、蔓の先端部は葉腋にできた腋芽である可能性を示唆している。右の写真では主腋芽と葉の間に副芽が形成されている。
An inflorescence of Hydrocotyle leucocephala is not formed at the axil of a leaf but rather in an opposite position. This suggests sympodial branching, in which an inflorescence is a terminal shoot and an extending vegetative shoot is an axillary shoot of a leaf. In the right picture, a secondary axillary shoot is formed between a main axillary shoot and a leaf.

チドメグサの場合も花序が頂生し、仮軸分枝によって茎が形成されている。
Sympodial branching is also observed in Hydrocotyle sibthorpioides.

オジギソウの葉は機械刺激、温度刺激、化学刺激、あるいは他の刺激によって、1秒以内に動く。この動きはお辞儀運動と呼ばれている。お辞儀運動はオジギソウ属で8回独立に進化した(Simon et al. 2011)。ただ、これら以外の種もオジギソウほど速くないが運動をするものがある(Simon et al. 2011)。お辞儀運動は以下の適応的意義があると考えられている。(1)捕食昆虫を驚かせたり振り落としたりする (Pickard 1973)、(2)食べごたえの少ない葉だと思わせる(Braam 2004)、(3)視界から消える(Eisner 1981)、(4)棘を表出する(Eisner 1981)。オジギソウと同じ属だが動かない種を隣あわせて植えると、動かない種の方がたくさん害虫に食べられてしまったという観察もある(Pickard 1973)。葉が回復するまでの時間は光合成に有効な光量が少ないほど速い(Jansen et al. 2011)。葉が閉じることは防御であるとすると、エネルギーに余裕のあるときは葉の閉じている時間を長くして防御に多くの投資をするが、エネルギーの少ないときは防御への投資を少なくしていると解釈されている(Jansen et al. 2011)。どんな分子機構でこのような仕組みを作っているのだろうか。
Leaves of Mimosa pudica move within a second by mechanistic, thermal, chemical, or other stimuli, which is called as seismonastic movement. Seismonasty evolved at least eight times in Mimosa (Simon et al. 2011). The sensitivity varies between species and multiple Mimosa species move more slowly in some extent (Simon et al. 2011). Seismonasty is proposed to be adaptive as defense strategy: (1) startling or throwing predator insects (Pickard 1973), (2) showing as less voluminous meals (Braam 2004), (3) disappearing from the view (Eisner 1981), and (4) pronounced increase in thorn exposure (Eisner 1981). When M. pudica and non-motile Mimosa species are planted side by side, the latter suffered more severely than the former did (Pickard 1973). Time to reopen leaves decreased in proportion to the decrease of photosynthetically active radiation (Jensen et al. 2011). This is interpreted that M. pudica accepts greater risk when energetically stressed as animals do (Jensen et al. 2011). How is such system operated at the molecular level?
Braam, J. (2005). In touch: plant responses to mechanical stimuli. New Phytol. 165, 373-389.
Eisner, T. (1981). Leaf folding in a sensitive plant: A defensive thorn-exposure mechanism? Proc. Natl. Acad. Sci. USA 78, 402-404.
Jensen, E.L., Dill, L.M., and Cahill, J.F., Jr. (2011). Applying behavioral-ecological theory to plant defense: light-dependent movement in Mimosa pudica suggests a trade-off between predation risk and energetic reward. Am Nat 177, 377-381.
Pickard, B.G. 1973. Action potentials in higher plants. Bot. Rev. 39: 172-201.
Simon, M.F., Grether, R., de Queiroz, L.P., Sarkinen, T.E., Dutra, V.F., and Hughes, C.E. (2011). The evolutionary history of Mimosa (Leguminosae): toward a phylogeny of the sensitive plants. Am. J. Bot. 98, 1201-1221.

When the leaf is touched,

視界から消える。
The leaf disappeared from the view.

Matricaria sp.の花序は開花当初は辺縁花の花弁が立ち上がり上方を向くが、花が咲き進むにつれ、下方を向くとともに、花托がせり上がり、種子散布に適している。円錐型にせり上がるのはどうしてだろう。どうやって成長が調和しているのだろうか。
Marticaria chamomilla (synonym: Matricaria recutita)はカモミールとしてハーブティーに用いられる。
The receptacle of Matricaria sp. Inflorescence grows up in a cone shape while flowering from proximal to distal parts. How is growth of cells in different parts coordinated to form the shape.
Marticaria chamomilla (synonym: Matricaria recutita) is chamomile used as herbal tea.

フラグミペディウムは新熱帯に分布し、パフィオペディラムは旧熱帯に分布する。Phragmipedium caudatumとPaphiopedilum sanderianumの花弁は50 cm程にまで伸長し、花粉媒介のために平行進化したと考えられる。伸長した花弁上には匂いを出す腺があり、ハナアブを呼び寄せ、花弁づたいに雄蕊雌蕊複合体へと誘導している(Dressler and Pupulin 2011)。どのように花弁を長くしたのだろうか。どうしてこれらの種だけ長い花弁を持ち得たのだろうか。
Phragmipedium is distributed in neo-tropics, while Paphiopedilum is in old tropics. Elongated petals evolved in both Phragmipedium caudatum and Paphiopedilum sanderianum in parallel for pollination. The elongated petals bear flagrance glands on the surface and hoverflies climb up to the stamen-gynoecium complex. How do these species change the petal morphology? Why could only these species change?
Dressler R.L. and Pupulin, F. 2011. The long-petaled tropical American lady’s slipper orchids. Lindleyana 26: 626-631.

Tank, D.C. and Donoghue, M.J. 2010. Phylogeny and phylogenetic nomenclature of the Campanulidae based on an expanded sample of genes and taxa. Syst. Bot. 35: 425-441.

フェリネ科はフェリネ属のみを含み、11種がニューカレドニアのみから知られている(Barriera et al. 2006)。
Phelline is monogenic in the Phellinaceae and11 species are known only in New Caledonia (Barriera et al. 2006).
Barriera, C., Savolainen, V., and Spichiger, R. 2006. Phellinaceae. Pp. 608-610, in Kadereit, J. W. and Jeffrey, C. (eds), The Families and Genera of Vascular Plants VIII Asterales. Springer, Berlin.

Phelline comosaの葉は偽輪生となる。
Leaves of Phelline comosa are clustered in tight spirals at the tip of branches.

葉序は互生だが、数枚ごとに葉と葉の間の茎(節間部)の伸長が長くなるために、輪生のように見える。
Leaves are alternate but some internodes are longer than other internodes.

Phelline comosa には成長期と休眠期がある。休眠期の茎頂を見ると、普通葉の後に鱗片葉が形成されている。ということは、成長期の終わりに鱗片葉を作るのだろう。そして、節間(白色矢印は鱗片葉の落ちた後)や節間遠位部にも鱗片葉があることから、休眠中の茎頂にすでに鱗片葉原基が形成されていて成長期初期にそれらがまず展開し、その後で、普通葉が展開するのか、あるいは、成長期初期に茎頂で最初に新しい鱗片葉原基が形成されるのだろう。Phelline comosaのフェノロジーについての報告は見つけられなかったが、年間を通して茎頂を切片による光学顕微鏡観察、走査電子顕微鏡観察すれば、季節変化と異型葉形成との関係がわかりそうである。
鱗片葉の適応的な意義はなんだろうか。休眠期に形成されることから、休眠中の茎頂保護に役立っているのかもしれない。多くの種子植物は休眠時に鱗片葉を形成する。これらの鱗片葉は次の成長期のために作られた若い葉原基や葉と茎頂を乾燥や昆虫の食害から保護する適応的意義がある。これらに加え、すらっと細く伸びるPhelline comosaの茎は周りの枝に当たる可能性が高く、鱗片葉が傷害からの保護の役割を持つかもしれない。
Phelline comosa has growth and dormant phases. At the dormant phase, scale leaves cover the shoot tip, indicating that Phelline comosa forms scale leaves at the late stage of the growth phase. Since scale leaves also locate on the middle and distal parts of internodes, growth of scale leaves formed at the later stage of the growth phase may be arrested and starts again when dormancy is released. Otherwise, scale leaves are formed at the shoot apex when growth starts, before the formation of normal leaves. I could not find any reports on the phenology of Phelline comosa. Observation of the shoot apex with microscopy for sections and scanning electron microscopy will reveal the relationship between phenology and heteroblasty.
What is the adaptive significance of scale leaves? They should function to protect the shoot meristem from dryness and harmful insects as scale leaves in other seed plants. Furthermore, since stems of Phelline comosa are slender and easily shaken by wind, the scale leaves may protect the shoot meristem from collisions to surrounding trees.

鱗片葉と普通葉を作る発生プログラムはかなり異なったものなのだろうか、あるいは、同じプログラムを使っているが鱗片葉の場合は途中でプログラムが止まっているのだろうか。同じ発生プログラムを途中でとめて、最終形態を変える機構は異時性と呼ばれる現象の範疇にあり、いろいろな生物で知られているが、その分子機構はよくわかっていない。
被子植物では、休眠芽から茎が伸び出す時、最初に展開する葉は通常葉とは、葉序や形態が異なることが多い。また、側芽が伸び出す時は、通常葉とは違った葉序と形態の葉が形成され、前出葉(前葉)と呼ばれる。しかし、休眠芽の発生様式の分子機構はほとんど明らかになっていない。Phelline comosaの鱗片葉も似た機構を使っているのだろうか。
It is unknown whether developmental programs to form scale and normal leaves are similar or different. They may use a similar developmental program, which is arrested at some point in the scale leaf development not to fully make the leaf blade. This is a kind of heterochrony, whose molecular mechanisms are not well known.
It is often observed in angiosperms that morphology and phyllotaxis of early leaves are different from those of later leaves when winter buds start to grow. Lateral branches usually have prophylls with different morphology and phyllotaxis to regular leaves. Molecular mechanisms to form such different types of leaves are mostly unknown. It is also unknown whether scale leaves of Phelline comosa use similar mechanisms.
成長期に入り茎が伸長する時、鱗片葉の節間は通常葉の節間よりも長く伸びる。鱗片葉の形成と茎の伸長には関係があるのだろうか。
Phelline comosa has growing and dormant phases. Just after the growing phase starts, internodes of scale leaves are extensively grow. Is there a relationship between scale leaves and rapid growth?

茎の伸長部周辺には鱗片葉がある。普通葉の節間は鱗片葉の節間よりも短い。
Internodes of scale leaves are longer than those of normal leaves.

休眠芽で形成されていた鱗片葉の節間長は通常葉の節間長と変わらない。
Internodes of scale leaves at the base of dormant bud are similar to those of normal leaves.

節間の途中に付く鱗片葉。
Some scale leaves locate on the longer internodes.

ツツジ科イチヤクソウ亜科 Pyroloideae の種も偽輪生になる。コイチヤクソウでは、Phelline comosaのように鱗片葉の付く位置で茎が伸長している。
Relationship between scale leaves and long internode is also observed in other plants. Orthilia secunda (Pyroloideae, Ericaceae) forms pseudo-verticillate leaves and internodes with scale leaves longer than those with normal leaves, as observed in Phelline comosa.

Pyrola chloranthaとイチゲイチヤクソウもツツジ科イチヤクソウ亜科に属する。Pyrola chloranthaもコイチヤクソウのように茎の伸長部に鱗片葉が形成されている。一方で、イチゲイチヤクソウの地上茎では鱗片葉が形成されても茎は伸長しない。Phelline comosa やイチヤクソウ亜科の鱗片葉と茎の伸長の関係は、異型葉性と節間伸長の関係、葉形態と茎形態との関係、長枝と短枝の関係などの植物発生学における一般的な問題につながっていると思われる。
Pyrola chlorantha and Moneses uniflora also belong to Pyroloideae in the Ericaceae. Scale leaves and internode growth are related in Pyrola chlorantha, while they are not in Moneses uniflora. The problem of the relationships between scale and normal leaves in Phelline comosa and Pyroloideae are related to the general questions on the relationships between heteroblasty and internode growth, between leaf and stem morphology, and between long and short branches.

フェリネ属は花序を腋生し、雌雄異株 (Barriera et al. 2006)。Phelline comosaの雌花は小さな雄蕊を形成し花粉も産出するが(白矢印)、花粉に粘性があるかはわからない。
Species in Phelline form axillary inflorescences with dioecious flowers (Barriera et al. 2006). Female flowers of Phelline comosa form both a gynoecium and four stamens, although fertility of the produced pollen are unknown.
Barriera, C., Savolainen, V., and Spichiger, R. 2006. Phellinaceae. Pp. 608-610, in Kadereit, J. W. and Jeffrey, C. (eds), The Families and Genera of Vascular Plants VIII Asterales. Springer, Berlin.

近縁のアルゴフィルム科やアルセウオスミア科では柱頭が広がり、花柱が明確に区別できるが、フェリネ科では柱頭が開かず花柱が不明瞭である。
Style is not clear in Phelline, while that is conspicuous in closely related families, Argophyllaceae and Alseuosmiaceae.


フェリネ属は核果を形成する。中果皮と内果皮は剥がれやすく、内果皮が種皮のように見える。
Phelline forms drupes. Mesocarp and endocarp are easily separated.


ストラスブルゲリア科は単系属であるイクセルバ属とストラスブルゲリア属からなる。Ixerba brexioidesはニュージーランド固有 (Schneider 2006)、Strasburgeria callianathaはニューカレドニア固有 (Dickison 2006) である。
The Strasburgeriaceae includes two monospecific genera, Ixerba and Strasburgeria. Ixerba brexioides and Strasburgeria calliantha are endemic in New Zealand and New Caledonia, respectively.
Dickison, W.C. 2006. Strasburgeriaceae. In Kubitzki, K. ed. The Families and Genera of Vascular Plants IX. Pp. 446-448.
Schneider, J.V. 2006. Ixerbaceae. In Kubitzki, K. ed. The Families and Genera of Vascular Plants IX. Pp. 205-207.

4枚の心皮が後天的合着する。
Four carpels are postgenitally fused.


ストラスブルゲリアは長さ6センチくらいの大きな果実を作る。リンゴに似た甘い匂いがする。
Strasburgeria calliantha forms a large fruit approximately 6 cm long with sweet smell similar to apple.

果実は落下後、腐って心皮の癒合面から割れ目が入る。4つの心皮には分かれやすい。しかし、それ以外の部分、例えば、心皮の背側はナイフで切ってもうまく切れないほど維管束由来の繊維が発達している。どうやって種子は発芽するのだろうと思ったら、子房の中心側からは種を取り出すことができた。
Fallen fruits are rotted and dehisced at the septicidal planes. It is easy to break the fruit at the septicidal planes, but is very difficult to cut at the loculicidal parts even with a knife because of massive fibers originated from vasculatures. Seeds are possible to be picked from the inner ridge of the quadrant fruit.

表皮と内部組織が腐っても維管束由来の繊維が1年以上残る。腐った果実は繊維のため、とても堅い。この自生地では発芽している種子は無かった。ストラスブルゲリアがどうしてこんなに大きくて甘い匂いを出す堅い実を付けるのかは謎である。Carpenter et al. (2003)はゾウに似て新大陸で絶滅したgomphotheresのような大型ほ乳類が果実を食べ、種子散布に寄与していたのではないかと考察している。
Epidermal and ground tissue of fruits are decayed but fibers remain more than one year. The decayed fruits are still very hard. Any seeds were not germinated in this locality. It is unknown why Strasburgeria form such large, sweet-smelled, and hard fruits. Carpenter et al. (2003) speculated that there were large mammals like gomphotheres (Janzen and Martin 1982) that ate the fruits and contributed to seed dispersal.
Carpenter, R.J., Read, J., and Jaffré, T. 2003. Reproductive traits of tropical rain-forest trees in New Caledonia. J. Tropical Ecol. 19: 351-365.
Janzen, D.H., and Martin, P.S. (1982). Neotropical anachronisms: the fruits the gomphotheres ate. Science 215, 19-27.


ウキクサ亜科は5属約35種がほぼ世界中に分布する (Landolt 1998)。サトイモ科の他の種に較べて姉妹種から分岐後の進化速度が速いが(Nauheimer et al. 2012)、理由は不明である。ウキクサは外群と較べ、形態変化が著しいが、外群からの分化時に、進化速度の速さゆえに、形態変化を起こし、その後、形態的には安定なので、進化速度が下がったのだろうか。
Lemnoideae contains five genera about 35 species, which are distributed worldwide (Landolt 1998). Evolutionary rate is accelerated in Lemnoideae (Nauheimer et al. 2012). Relationship between the accelerated evolutionary rate and conspicuous morphological differences from sister taxa is unknown.
Landolt, E. 1998. Lemanceae. In K. Kubitzki ed. The Families and Genera of Vascular Plants. Pp. 264-270.
Nauheimer, L., Metzler, D., Renner, S.S. 2012. Global history of the ancient monocot family Araceae inferred with models accounting for past continental positions and previous ranges based on fossils. New Phytol. 195: 938-950.


ウキクサ属、アオウキクサ属、ミジンコウキクサ属のフロンド形成過程の比較から、Lemon and Posluszny (2000)はフロンドが茎と葉の発生過程を混ぜたようなものだという従来の見解を支持している。Wolffia microscopicaのさらに詳細なフロンド発生過程(Sree et al. 2000)を勘案すると、フロンドの初期過程は葉原基の無い茎頂分裂組織のようで成長につれ基部側に新しい分裂組織を切り出しているようにも見える。そして、しばらくすると分裂組織を生み出さず葉として発生する。この発生様式はカワゴケソウ科のカワゴケソウやカワゴロモに似ている。これらの植物は、葉状器官の初期原基では茎頂分裂組織で発現するSTMやWUSが検出され、原基が大きくなると葉原基で発現するARPが検出される(Katayama et al. 2010)。ウキクサ亜科においてこれらの遺伝子や、茎頂分裂組織と葉原基の区別に関わるオーキシン動態を解析すると、フロンドと他の植物の茎葉との相同性が明らかになるのではないだろうか。
Based on the comparison of development of Spirodela, Lemna, and Wolffia, Lemon and Posluszny (2000) agreed to previous ideas that the frond is a developmental hybrid of leaf and stem origin. When the more detailed development of Wolffia microscopica (Sree et al. 2000) is incorporeted, it appears for me that the early development of the frond is shoot-like with producing detached meristems as daughter fronds and that the later development is leaf-like. This developmental process looks similar to that of the Hydrobryum japonicum and Cladopus doianus in the Posdostemaceae, in which the shoot meristem genes, STM and WUS are expressed at the early primordium and then are substituted by the leaf gene ARP in the later development (Katayama et al. 2010). Analyses of these genes as well as auxin which is involved in the differentiation of shoot meristem and leaves will give insight in the homology of the frond in Lemnoideae plants.
Katayama, N., Koi, S. and Kato, M. (2010) Expression of SHOOT MERISTEMLESS, WUSCHEL, and ASYMMETRIC LEAVES1 homologs in the shoots of Podostemaceae: Implications for the evolution of novel shoot organogenesis. Plant Cell 22: 2131-2140.
Lemon, G.D. and Posluszny, U. 2000. Comparative Shoot Development and Evolution in the Lemnaceae. Int. J. Plant Sci. 161: 733-748.
Sree, K.S., Maheshwari, S.C., Boka, K., Khurana, J.P., Keresztes, A., and Appenroth, K.-J. 2015. The duckweek Wolffia microscopica: a unique aquatic monocot. Flora 210: 31-39.

従来、 ウキクサ亜科は、1つの雌花と1つの雄花からなる花序を持つボタンウキクサと姉妹関係にあると考えられていたので、ウキクサ亜科の生殖器官は、雄しべのみからなる雄花と雌蕊のみからなる雌花を総苞が被っていると考えられてきた。しかし、ボタンウキクサとは姉妹関係に無いことがわかったので、現在では、花被を持たない雄蕊と雌蕊のみからなる1つの花が、1枚の葉的器官に被われていると解釈されている (Londolt 1998)。この葉的器官は、サトイモ科の他の種の仏縁苞と相同であるか、あるいは、花被に相同なのかはわからない。
The reproductive organs of Lemnoideae have been interpreted as an inflorescence with postulating a close relationship to Pistia. However, molecular phylogeny revealed that Pistia and Lemnoideae are not closely related. Now, one reproductive unit including one or two stamens and one pistil is interpreted as a flower.
A flower without petals and petals is subtended by a membranous scale like leaf. It is unknown whether this scale is a tepal or a spathe. Here it is tentatively indicated as a spathe.


イボウキクサは2本の雄蕊と1本の雌蕊を形成する。
Lemna gibba forms two stamens and one gynoecium.

ミジンコウキクサ属は1本の雄蕊と1本の雌蕊を形成する。雄蕊は200 µmほど、雌蕊は100 µmほどである。フロンドから雌蕊が突出し、次に、雄蕊が突出する。雄蕊の基部に前に出た雌蕊が残っている。漏斗状の柱頭から分泌滴が出て受粉に機能する。Wolffia microscopicaは他のミジンコウキクサの種よりも頻繁に花を形成するので花を必要とする実験に適している。
Wolffia forms a single stamen and a gynoecium. The lengths of a stamen and a gynoecium are 200 µm and 100 µm, respectively.A gynoecium comes out and then a stamen does. At the base of the stamen, the gynoecium still remains. Exudate is formed at the tip of a funnel-like stigma for pollination. In comparison to other Wolffia species, W. microscopica frequently blooms and is useful for experiments in flowering.
Sree, K.S., Maheshwari, S.C., Boka, K., Khurana, J.P., Keresztes, A., and Appenroth, K.-J. 2014. The duckweed Wolffia microscopica: A unique aquatic monocot. Flora 210: 31-39.
イボウキクサ、Wolffia microscopicaの観察は京都大学の小山時隆先生に御教示いただきました。御礼申し上げます。
I appreciate Dr. Tokitaka Oyama for providing materials of Lemna gibba and Wolffia microscopica and kind instructions for the cultivation and flowering.

Seberg, O., Petersen, G., Davis, J. I., Pires, J. C., Stevenson, D. W., Chase, M. W., Fay, M. F., Devey, D. S., Jørgensen, T., Sytsma, K. J., and Pillon, Y. 2012. Phylogeny of the Asparagales based on three plastid and two mitochondrial genes. American J. Bot. 99: 875-889.
Steele, P. R., Hertweck, K. L., Mayfield, D., McKain, M. R., Leebens-Mack, J., and Pires, J. C. 2012. Quality and quantity of data recovered from massively parallel sequencing: Examples in Asparagales and Poaceae. American J. Bot. 99: 330348.

キジカクシ亜科は3個以上の種子を含む液果を形成し、ファイトメランのある種子を形成する。
Asparagoideae form berries with more than three seeds with phytomelan..
Rudall, P.J., Conran, J.G., and Chase, M.W. 2000. Systematics of Ruscaceae/Convallariaceae: a combined morphological and molecular investigation. Bot. J. Linn. Soc. 134: 73-92.


Eriospermumの全ての写真はSpecies Nurseryの藤川史雄様のご厚意によって藤川様の栽培品を撮影させていただきました。また、学名や記載内容について同好の方からご教示いただきました。この場を借りて感謝申し上げます。
約100種が南アフリカを中心に分布している (Perry 1994)。このうち約13種は葉の向軸側に突起を形成する。突起を形成する種のほとんどは同じ核型を持つことから単系統群であろうと考えられている(Vosa and Perry 2012)。
All photos of Eriospermum are took in Species Nursery by great courtesy of Mr. Fumio Fujikawa. I also appreciate an anonymous reader for suggestion on some scientific names.
Approximately 100 species are distributed mainly in South Africa (Perry 1994). About 13 species form leaf appendages but others do not (Perry 1984). These appendaged species share similar karyotype and are inferred to be monophyletic (Vosa and Perry 2012).
Perry, P. L. 1994. A revision of the genus Eriosperma (Eriospermaceae). Contrib. Bolus Herb. 17: 1-320.
Vosa, C.G. and Perry, P.L. 2012. On the cytotaxonomy of the genus Eriospermum Jacq. Ex Willd. (Erisspermaceae). Caryologia 52: 117-125.

Eriospermumの根茎は胚軸起源である(Perry 1994)。
The corm of Eriospermum is originated from a hypocotyl (Perry 1994).

Eriospermumの種には、変わった花序や突起のある葉を持つものがある。E. bayeriは花後も花序が長く残存し、光合成を行う。E. aphyllumは花を付けない栄養性花序様器官を持つ。光合成に役立っていると考えられる。E. dregeiは複雑に分岐して毛がたくさん生えた栄養性花序様器官を形成する。光合成とともに、霧がかかるような乾燥地帯に分布していることから、霧から水分を選る役割があるのかもしれない。E. cervicorneは葉の上に多数の突起がある。複雑に分岐した栄養性花序様器官が苞葉に癒合しているように見える。E. alcicorneやE. appendiculatumは葉の上に突起を形成する。これらの突起も、栄養性花序様器官が癒合してできたのかもしれない。発生過程や突起の維管束の道管と師管の位置関係を調べると、突起が花序起源かどうかわかるかもしれない。また、他の植物の花序形成に関わる遺伝子が発現しているか、発現しているならどう変わっているかも調べると面白そう。
注:ここでは平面葉以外の立体的なシュート的器官や葉状突起を、花序が変形してできた器官ではないかという仮説のもと「栄養性花序様器官」と呼んでいますが、本当に花序由来かはまだわかりません。今後の研究で明らかになっていくと思います。
Some species of Eriospermum has unusual inflorescence-like organs and leaf appendages. E. bayeri has long-lasting inflorescence after blooming. E. aphyllum forms non-reproductive inflorescence-like organ, which is formed without flowers and stout stems. E. dregei forms multiply divided vegetative inflorescence-like organ with dense hair. A leaf of E. cervicorne has appendages, which appear to be a fusion of multiply divided vegetative inflorescence-like organ. E. alcicome and E. appenduculatum has appendages on their leaves. These appendages may be homologous to branches of a vegetative inflorescence-like organ. Development and anatomical analyses of these appendages will give insight on the origin of leaf appendages in Eriospermum.
The unusual vegetative inflorescence-like organs and leaf appendages appear to be adaptive for dry environments to capture water from fogs.
The number and size of appendages are variable in a population of some species, but it appears that the total area of appendages and leaf is mostly constant between leaves (Perry 1994). This suggests that appendages are useful for photosynthesis.
Perry, P. L. 1994. A revision of the genus Eriosperma (Eriospermaceae). Contrib. Bolus Herb. 17: 1-320.


E. porhyrovalveの花序は花後枯れる。
Inflorescences of E. porhyrovalve is senesced when seeds are not harvested.

E. cooperiの花序は花後枯れる。
Inflorescences of E. cooperi is senesced when seeds are not harvested.

Eriospermum bayeriの花序は種子が稔らなくても残存し、光合成をしているようだ。
Inflorescences of Eriospermum bayeri is alive even after no fruits are formed.

Eriospermum aphyllumの花序様の器官は花を付けず、長く維持されることから、栄養性の花序かもしれない。トウダイグサ科の植物のように、葉でなく柱状の茎を持つことは、蒸散を防ぎ、乾燥環境に適応していると考えられる。
Eriospermum aphyllum forms a vegetative inflorescence-like organ, which does not have any flowers and is long-lasting. As some succulent species in Euphorbiaceae, the vegetative inflorescence-like organ should be adaptive under dry conditions.

Eriospermum paradoxumは複雑に分岐する栄養性花序様器官を形成する。枝分かれし、毛が生えた栄養性花序様器官は乾燥地域で、霧から水分をとるのに役立っていると考えられる。
Eriospermum paradoxum forms a multiply divided vegetative inflorescence-like organ. Hairy and branched vegetative inflorescence-like organs should be adaptive for dry conditions to capture water from fogs (Vogel and Müller-Doblies 2011).
Vogel, S. and Müller-Doblies, U. 2011. Desert geophytes under dew and fog: The “curly-whirlies” of Namaqualand (South Africa). Flora 206: 3-31.

やや不規則な総状花序のような分岐の栄養性花序様器官を形成する。
Partly disarranged raceme-like vegetative inflorescence-like organs.

複雑に分岐し毛が生えた栄養性花序様器官。栄養性花序様器官を形成しない葉は両面に毛が生えているが、栄養性花序様器官を抱く葉(苞)は向軸側に毛が無い。どうしてなのだろうか。
Complex, branched and hairy vegetative inflorescence-organs. A leaf is hairy on both sides but the adaxial surface of a bract is relatively glabrous. Why?

複雑に分岐し毛が生えた栄養性花序様器官。複雑な分岐で、どんな発生的な規則でできているのか不明。
Complex, branched and hairy vegetative inflorescence-like organs. The pattern of branching is complicated and what is a developmental rule to organize the form..

分岐した栄養性花序様器官が苞に癒合しているように見える。
A vegetative inflorescence-like organ appears to be fused with a bract.

分岐した栄養性花序様器官が苞に癒合しているように見える。
A vegetative inflorescence-like organ appears to be fused with a bract.

分岐した栄養性花序様器官が苞に癒合しているように見える。
A vegetative inflorescence-like organ appears to be fused with a bract.

葉の上の突起は葉状。栄養性花序様器官が癒合し、枝が葉状化したのだろうか。
Appendages are leaf-like. After the fusion of a vegetative inflorescence-like organ, branches may be changed to be leaf-like.

栄養花序様器官が癒合したなら、突起の出ている部分から葉の基部にわたり、維管束が本来の苞の維管束と重複しているかもしれない。
If vegetative inflorescence-like organ is fused, vascular bundles may be doubled from the junction to a leaf base.

E. appendiculatumの突起は栄養性花序様器官が癒合したものなのだろうか。突起の先端の毛が葉縁の毛と似ているので、もしかすると、他の種類の突起とは違って、葉縁で発現すべき遺伝子が異所的に発現することで葉の表面の細胞が葉縁として運命づけられたのかもしれない。
It is quite hard to say these appendages are homologous to the vegetative inflorescence-like organ. Hair at the tip of an appendage is similar to that at the leaf margin, suggesting that the appendage captures a part of fate of leaf margin with meristematic activity.












{kind=link}