プレイオスタキア属 Pleiostachya Pleiostachya pruinosa_1: Cropped a photo by Wayne A. Hughes, https://www.inaturalist.org/photos/149456900, CC BY-NC Pleiostachya pruinosa-2: Cropped a photo by Kevin Faccenda, https://www.inaturalist.org/photos/550661109, CC BY Pleiostachya leiostachya-1: Cropped a photo by Marvin López M. , https://www.inaturalist.org/photos/43078880, CC BY-NC Pleiostachya leiostachya-2: Cropped a photo by Marvin López M. , https://www.inaturalist.org/photos/43026397, CC BY-NC
The genus Pleiostachya is distributed from Central America to northern South America and comprises three recognized species (POWO). Pleiostachya pruinosa has almost glabrous inflorescence bracts, whereas in P. leiostachya the bracts are densely villous, and the two species also differ in the colour of the abaxial leaf surface (Hammel 1986).
プレイオスタキア属の特徴 Characteristics of the genus Pleiostachya Ischnosiphon petiolatus: cropped a photo by Pierrot F., https://www.inaturalist.org/photos/617327689, CC BY-NC
プレイオスタキア属の特徴 Characteristics of the genus Pleiostachya
The genus Pleiostachya differs from its sister genus Ischnosiphon in forming laterally flattened inflorescences (Andersson 1998).
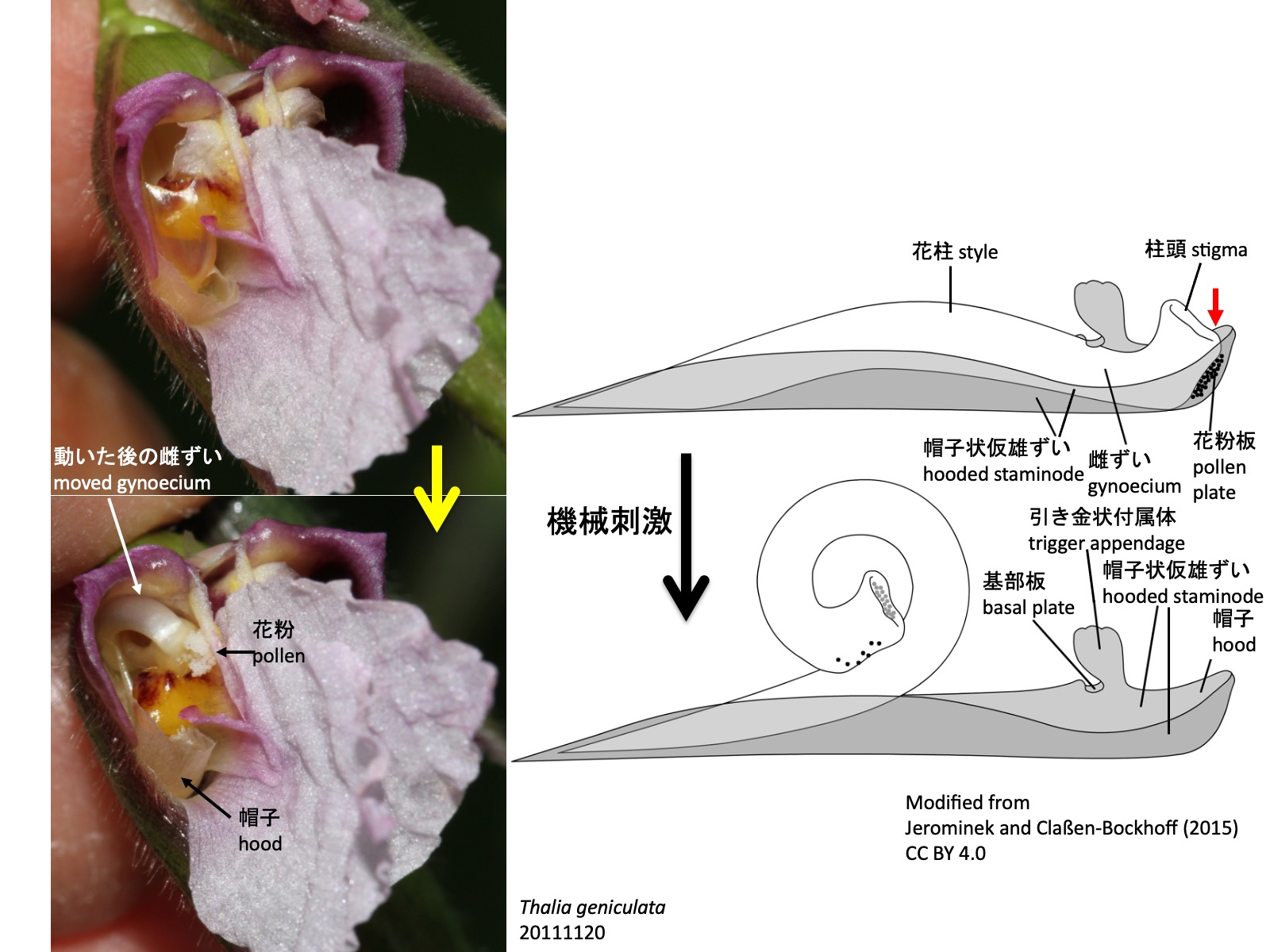
クズウコン科における花柱運動と二次花粉提示 Style movement and secondary pollen presentation in Marantaceae Jerominek, M., and Claßen-Bockhoff, R. (2015). Electrical signals in prayer plants (Marantaceae)? insights into the trigger mechanism of the explosive style movement. PLoS One 10, e0126411. https://doi.org/10.1371/JOURNAL.PONE.0126411. CC BY 4.0.
クズウコン科における花柱運動と二次花粉提示Style movement and secondary pollen presentation in Marantaceae
多くの種では、花粉板と柱頭の間が帽子状仮雄ずいの先端部によってしっかり閉じられているが(模式図の赤色矢印)、Pleiostachya pruinosaではこの密着が緩く、蕾の中で花粉が柱頭に漏れ出し、自家受粉する場合がある (Ley and Claßen-Bockhoff 2012)。このため、温室など送粉昆虫のいない環境でも種子ができやすい。
In flowers of Marantaceae, mechanical stimulation causes the style to move, resulting in secondary pollen presentation. In the bud, the distal hood of the hooded staminode encloses the distal part of the pistil, including the dorsal side of the stigma, together with the distal part of the fertile stamen, namely the anther. Because the style elongates more than the hooded staminode, it grows in a backward-bent position and mechanical tension is generated. The hooded staminode bears a projection that forms the trigger appendage and the connected basal plate. When a pollinating insect touches the trigger appendage, the basal plate that holds the tensioned style in place is displaced, and the style curls adaxially. At this moment, the style moves with pollen attached to the pollen plate on the dorsal side of the stigma; as a result, the pollen is presented at the front of the flower and deposited on the pollinating insect. In many species, the passage between the pollen plate and the stigma is tightly closed by the distal part of the hooded staminode (red arrow in the diagram). In Pleiostachya pruinosa, however, this contact is looser, allowing pollen to enter the stigma in the bud and sometimes resulting in self-pollination (Ley and Claßen-Bockhoff 2012). This may explain why seeds are readily produced even in environments without pollinating insects, such as greenhouses.
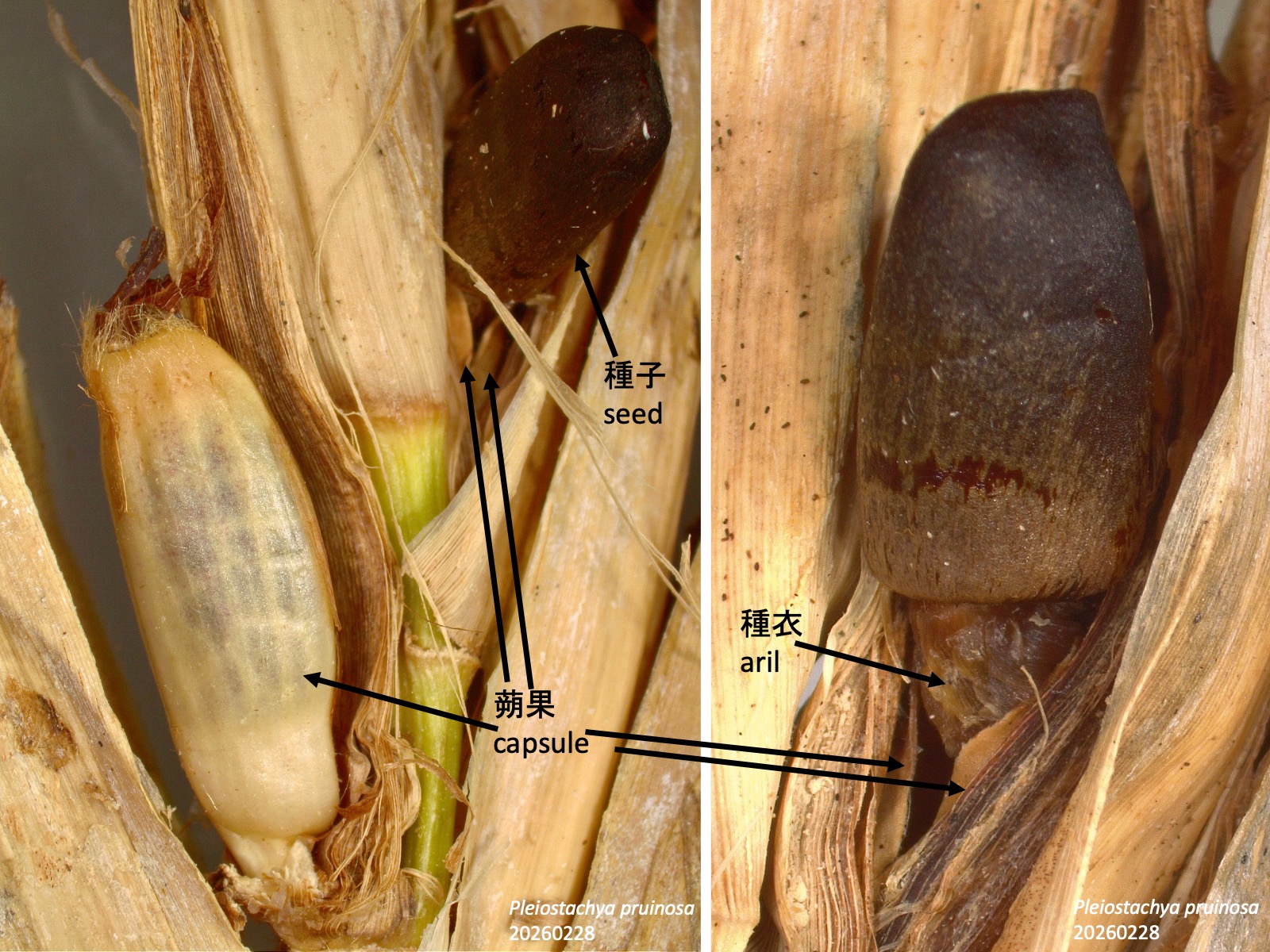
クズウコン科の蒴果から押し出される種子Seeds protruding from capsules in Marantaceae
In Marantaceae, when the seeds mature within the capsule, the aril develops and expands, causing the fruit wall to dehisce and the seeds to be pushed outward (Grootjen 1983). In Pleiostachya pruinosa, black seeds become exposed as they protrude from pale brown, senescing capsules (yellow arrow). How are these exposed seeds dispersed?
クズウコン科の種子散布-1 Seed dispersal in Marantaceae_1 Calathea lutea: cropped a photo by Oscar Johnson, https://www.inaturalist.org/photos/209790037, CC BY-NC-ND Calathea inocephala: cropped a photo by Vanesa Cruz Jumbo, https://www.inaturalist.org/photos/205659457, CC BY-NCクズウコン科の種子散布-2 Seed dispersal in Marantaceae_2 Pleiostachya leiostachya: Cropped a photo by Eduardo Chacón Madrigal, https://www.inaturalist.org/photos/80935719, CC BY Goeppertia marantifolia: Cropped a photo by Marco Acuña, https://www.inaturalist.org/photos/324684511, CC BY-NC
クズウコン科の種子散布 Seed dispersal in Marantaceae
クズウコン科Marantaceaeの種には、鳥散布、アリ散布、あるいはその両方による散布が知られている (Horvitz 1991; Horvitz et al. 2002)。コスタリカでは、クズウコン科の数種について、鳥が種子を果序から持ち去る割合、および種子を落葉層の上に置いた場合にアリ類が持ち去る割合を調べる研究が行われた (Horvitz et al. 2002)。
Calathea lutea、Calatheainocephala、Pleiostachya pruinosa、Goeppertiamarantifoliaでは、それぞれ異なった形態の種子が蒴果から露出する。C. luteaでは桃色の蒴果から橙色の種皮をもつ種子が、C. inocephalaではオレンジ色の蝋質の蒴果から青色の種子が露出する。P. pruinosaでは薄茶色の蒴果から黒色の種子が露出し、G. marantifoliaでは青色の種子が露出する。なお、上の写真は P. leiostachyaである。
In Marantaceae, seed dispersal by birds, by ants, or by both is known in several species (Horvitz 1991; Horvitz et al. 2002). In Costa Rica, a study examined, for several species of Marantaceae, the proportion of seeds removed by birds from infructescences and the proportion of seeds removed by ants when the seeds were placed on the leaf litter (Horvitz et al. 2002).
In Calathea lutea, Calathea inocephala, Pleiostachya pruinosa, and Goeppertia marantifolia, seeds with different morphologies are exposed from the capsules. In C. lutea, seeds with orange arils are exposed from pink capsules, whereas in C. inocephala, blue seeds are exposed from orange, waxy capsules. In P. pruinosa, black seeds are exposed from pale brown capsules, and in G. marantifolia, blue seeds are exposed. The photograph above shows P. leiostachya.
The proportion of seeds removed from infructescences by birds was approximately 80% in C. lutea, 40% in C. inocephala, 30% in P. pruinosa, and 10% in G. marantifolia. In contrast, when seeds were placed on the leaf litter, the proportion removed by ants was nearly 100% in C. lutea, nearly 0% in C. inocephala, approximately 40% in P. pruinosa, and approximately 90% in G. marantifolia. However, the data for C. inocephala were obtained at a different site. Calathea inocephala has larger seeds than the other species, and this may explain why ants did not remove them. In the other species, seeds may be dispersed by birds while they remain attached to the infructescence and by ants after they fall from the infructescence, although the degree of dependence on each disperser appears to vary among species and habitats.
謝辞 Acknowledgements
本稿執筆にあたり、名古屋市東山動植物園の栽培株を観察させていただきました。心より感謝申し上げます。
I am sincerely grateful to the Nagoya Higashiyama Zoo & Botanical Gardens for allowing me to observe cultivated plants for the preparation of this article.
引用文献 References
Andersson, L. (1998) Marantaceae. In K. Kubitzki (ed.) 1998. The Families and Genera of Vascular Plants. Vol. IV. Monocotyledons: Alismatanae and Commelinanae (except Gramineae). Springer. Berlin. Pp. 278-293.
Grootjen, C.J. (1983). Development of ovule and seed in Marantaceae. Acta Bot. Neerl. 32, 69–86.
Hammel, B.E. (1986). The vascular flora of La Selva biological station, Costa Rica Marantaceae. Selbyana 9, 234–242.
Holvitz, C.C. 1991. Light environments, stage structures and dispersal syndromes of Costa Rican Marantaceae. In Huxley, C.R. and Cutler, D.F. eds, Ant-Plant Interactions. Oxford Univ. Press. pp. 463-485.
Holvitz, C.C., Pizo, M.A., y Belllo, B.B., LeCorff, J. and Dirzo, R. 2002. Are plant species that need gaps for recruitment more attractive to seed-dispersing birds and ants than other species? In Levey, D.J, Silva, W.R., and Galetti, M. eds, Seed Dispersal and Frugivory: Ecology, Evolution and Conservation. CABI Publishing, Oxon, UK. pp. 145-159.
Jerominek, M., and Claßen-Bockhoff, R. (2015). Electrical signals in prayer plants (Marantaceae)? insights into the trigger mechanism of the explosive style movement. PLoS One 10, e0126411. https://doi.org/10.1371/JOURNAL.PONE.0126411.
Ley, A.C., and Claßen-Bockhoff, R. (2012). Floral synorganization and its influence on mechanical isolation and autogamy in Marantaceae. Bot. J. Linn.Soc. 168, 300–322. https://doi.org/10.1111/J.1095-8339.2011.01202.X.
Genlisea is a genus of carnivorous plants in the family Lentibulariaceae, native to South America and central to southern Africa (Fleischmann 2012). The photographs show Genlisea roraimensis growing in a wetland on the summit of Mount Roraima in the Guiana Highlands of Venezuela. Although the aboveground morphology resembles that of its sister genus Utricularia (left photograph and upper right photograph), the underground morphology differs markedly (lower right photograph).
ゲンリセア属の種では根が発生せず、地下に捕虫葉が伸長する。
Species of Genlisea do not develop roots; instead, they produce underground trapping leaves.
捕虫葉は以下のように発生する (Lloyd 1942)。図(1)は葉原基を側面からみた図、それ以外は葉の表側(向軸側)から見た図である。(1) 葉原基の先端に窪みが生じる。葉原基の伸長に伴ってこの窪みは深くなり、やがて筒状になる。サラセニア プルプレアやイトタヌキモの嚢状葉のできかたから考えると(Fukushima et al. 2015; Whitewoods et al. 2020)、筒の外側が平面葉の表側(向軸側)、筒の内側が表側(背軸側)に相当し、表側と裏側の境界領域が伸長していると推定される。 (2) 葉原基の先端部が左右にやや広がり始める。 (3) さらに、パイプを押しつぶしたような状態で左右に伸長する。捕虫嚢もさらに膨らむ。(4) 先端部は押しつぶされたような状態のままさらに左右に広がり、頸部は伸長し、捕虫嚢もさらに膨らむ。(5) 先端部はさらに左右に伸長して二叉となり、腕部と呼ばれる。二叉部分は押しつぶされたような状態で、筒の表側部分と裏側部分が重なり、二重になっている。
The trapping leaf develops as follows (Lloyd 1942). Figure (1) shows a leaf primordium in lateral view, whereas the other figures show the leaf from the upper, or adaxial, side. (1) A depression forms at the tip of the leaf primordium. As the leaf primordium elongates, this depression becomes deeper and eventually forms a tube. Based on the developmental processes of pitcher-like leaves in Sarracenia purpurea and Utricularia gibba (Fukushima et al. 2015; Whitewoods et al. 2020), the outer side of the tube is inferred to correspond to the upper, or adaxial, side of a flat leaf, and the inner side to the lower, or abaxial, side; the boundary region between the upper and lower sides is therefore inferred to elongate. (2) The distal part of the leaf primordium begins to expand slightly to the left and right. (3) It then elongates laterally, as if a pipe had been flattened. The trap bladder also swells further. (4) The distal part continues to expand laterally while remaining flattened, the neck elongates, and the trap bladder continues to swell. (5) The distal part elongates further to the left and right and becomes bifurcated, forming the structures known as arms. The bifurcated region remains flattened, with the upper and lower portions of the tube overlapping to form a double-layered structure.
The inner side of the arm is thought to correspond to the upper, or adaxial, side of a flat leaf, whereas the outer side corresponds to the lower, or abaxial, side. Thus, the arm can be interpreted as a structure in which the leaf is valley-folded toward the upper side. In the figure above, the dark gray region indicates the inner side of the arm, and the light gray region indicates the outer side.
From the epidermal cells on one side of the folded structure, hairs called bridging trichomes (Juniper et al. 1989), which are derived from epidermal cells, elongate. Like the other hairs formed on the trapping leaf, each bridging trichome consists of three cells: a base cell, a middle cell, and an end cell (Lloyd 1942). Several types of hairs with the same basic structure are formed on the trapping leaves of Genlisea, but different names have been used for these three cells depending on the hair type (Lloyd 1942; Fleischmann 2012). For example, the middle cell of a bridging trichome has been called a prop cell (Lloyd 1942; Fleischmann 2012). Here, to clarify homology, these cells are referred to as the base cell, middle cell, and end cell in all hair types.
The bridging trichomes adhere to epidermal cells on the opposing surface, thereby forming regularly spaced gaps inside the folded arm. As the arm elongates, it twists in the direction indicated by the arrows in the figure.
As the arm twists in the direction indicated in the preceding figure, it eventually assumes the form shown in the upper left figure. The photograph on the right is a scanning electron micrograph of an arm at the folded stage, before adhesion by the bridging trichomes, opened and viewed from the inside. One prospective adhesion site of a bridging trichome is indicated. On the inner surface, which corresponds to the upper side of the leaf, detentive hairs (Juniper et al. 1989; Fleischmann 2012; trapping hairs in Lloyd 1942) are arranged in rows directed toward the trap bladder. When prey enters through the gaps in the arm, these detentive hairs guide it toward the trap bladder.
引用文献 References
Fleischmann, A. (2012). Monograph of the Genus Genlisea (Redfern Natural History Productions).
Fukushima, K., Fujita, H., Yamaguchi, T., Kawaguchi, M., Tsukaya, H., and Hasebe, M. (2015). Oriented cell division shapes carnivorous pitcher leaves of Sarracenia purpurea. Nat. Commun. 6, 6450.
Lloyd, F.E. (1942). The Carnivorous Plants (Dover Publications, Inc.).
Whitewoods, C.D., Whitewoods, C.D., Gonçalves, B., Cheng, J., Cui, M., and Kennaway, R. (2020). Evolution of carnivorous traps from planar leaves through simple shifts in gene expression. Science (1979). 367, 91–96.
カテゴリー:未分類|ゲンリセア属の捕虫葉の発生 Development of the Trapping Leaves in Genlisea はコメントを受け付けていません
Gloriosa belongs to Colchicaceae and comprises 11 known species, distributed mainly in Africa and Asia (POWO).
Gloriosa rigidifolia: Cropped from a photo by Manuel R Popp, https://www.inaturalist.org/photos/174698901, CC BY Gloriosa rigidifolia: Cropped from a photo by Manuel R Popp, https://www.inaturalist.org/photos/169349621, CC BY
Gloriosaには花被片が強く反り返り、花柱がほぼ直角に曲がる種(例えばGloriosa superba)と、花被片が強く反り返らず、花柱がほとんど曲がらない種(例えばGloriosa rigidifolia)があり、後者はかつてリットニア属Littoniaとして扱われたこともあったが、分子系統解析によりグロリオサ属と単系統群を形成することがわかり、現在ではグロリオサ属に統合されている (Thi et al. 2013)。
Gloriosa includes species with strongly reflexed tepals and a style bent almost at a right angle, such as Gloriosa superba, as well as species with tepals that are not strongly reflexed and a style that is scarcely bent, such as Gloriosa rigidifolia. The latter were once treated as members of the genus Littonia, but molecular phylogenetic analyses showed that they form a monophyletic group with Gloriosa. They are therefore now included in Gloriosa (Thi et al. 2013).
グロリオサのほぼ直角に曲がる雌ずい a style of Gloriosa superba bent almost at a right angle
グロリオサのほぼ直角に曲がる雌ずい
a style of Gloriosa superba bent almost at a right angle
Gloriosa superbaの花柱は、発生過程で基部が向軸側(花の上側)へほぼ直角に屈曲する。子房では向軸側の子房室の大きさが背軸側の子房室よりも小さく、蕾の段階ですでに屈曲が認められる。柱頭は葯より突出するため、他個体の花粉を付着した送粉昆虫であるシロチョウ科のチョウ類は、訪花時にまず柱頭に接触しやすい。その結果、自家受粉の割合が低下し、他家受粉の割合が高まると考えられる(Daniels et al. 2020)。同様の雌ずいの屈曲は、Lilium martagonでも観察されている(Brantjes and Bos 1980, Corbera et al. 2018)。
In Gloriosa superba, the style becomes bent almost at a right angle toward the adaxial side, i.e. the upper side of the flower, during development. In the ovary, the adaxial locule is smaller than the abaxial locule, and the curvature is already evident at the bud stage. Because the stigma protrudes beyond the anthers, pierid butterflies carrying pollen from other individuals are likely to contact the stigma first when visiting the flower. This is thought to reduce the frequency of self-pollination and increase the frequency of cross-pollination (Daniels et al. 2020). A similar curvature of the pistil has also been observed in Lilium martagon (Brantjes and Bos 1980; Corbera et al. 2018).
グロリオサの長い口吻用の蜜腺 nectaries adapted for long proboscides in Gloriosa superba
グロリオサの長い口吻用の蜜腺
nectaries adapted for long proboscides in Gloriosa superba
ユリ目の共有派生形質は、花被片上に花被蜜腺を持つことである。Gloriosa superbaの蜜腺は花被片の向軸側基部にあり、チョウ類は左の写真の花弁向軸側の入口から口吻を差し込み(点線)、花被片基部にある蜜腺から吸蜜する。蜜腺への入口は、口吻の短い昆虫の吸蜜を制限し、口吻の長いチョウ類による訪花を促進していると考えられる(Daniels et al. 2020)。チョウ類が反り返った花被片の基部にある蜜腺へアプローチすると、翅の下面が葯に接触し、花粉が付着する(Daniels et al. 2020)。 A shared derived character of Liliales is the presence of tepal nectaries on the tepals. In Gloriosa superba, the nectaries are located at the adaxial base of the tepals. Butterflies insert their proboscides through the entrance on the adaxial side of the tepal shown in the photograph on the left (dotted line) and feed on nectar from the nectaries at the tepal base. The entrance to the nectary is thought to restrict nectar access by insects with short proboscides and to promote visitation by long-proboscid butterflies (Daniels et al. 2020). When butterflies approach the nectaries at the base of the reflexed tepals, the undersides of their wings come into contact with the anthers and become dusted with pollen (Daniels et al. 2020).
グロリオサの花被の色彩 Perianth coloration in Gloriosa
グロリオサの花被の色彩
Perianth coloration in Gloriosa
グロリオサの花被片に見られるオレンジ色から赤色と黄色の模様は、チョウ類に認識されやすく(Weiss 1997)、V字型の模様は蜜腺への誘引路として機能している可能性がある(Daniels et al. 2020)。 The orange-to-red and yellow pattern seen on the tepals of Gloriosa is easily recognized by butterflies (Weiss 1997), and the V-shaped pattern may function as a guide leading them to the nectaries (Daniels et al. 2020).
グロリオサの葉先の巻きひげ Leaf-tip tendrils of Gloriosa superba
グロリオサの葉先の巻きひげ
Leaf-tip tendrils of Gloriosa superba
グロリオサの葉身部の先端側 (写真の黒色矢印) では表裏の区別が明瞭である。一方、巻きひげとなる葉先部(赤色矢印)は、葉身部と異なり、やや太く細長い円柱状となる。このため外形は放射相称的に見えるが、維管束は背腹性を保持している (Arber 1920)。巻きひげの先端は硬く棘状となり、他物に引っかかりやすい。葉先は、他物に接触するかどうかにかかわらず、伸長を続ける。 In the distal part of the leaf blade of Gloriosasuperba (black arrow in the photograph), the distinction between the adaxial and abaxial surfaces is clear. In contrast, the leaf-tip region that develops into a tendril (red arrow) differs from the leaf blade in becoming a somewhat thickened, slender cylindrical structure. Thus, its external morphology appears radially symmetrical, but the vascular bundles retain dorsiventrality (Arber 1920). The tip of the tendril becomes hard and spine-like, making it easy to catch on surrounding objects. The leaf tip continues to elongate regardless of whether it comes into contact with surrounding objects.
Plants of Colchicaceae form corms or rhizomes (Nordenstam 1998). The axillary buds of the first two leaves of the current-year shoot become the shoot apices of the following year’s shoots (Nordenstam 1998). Because the current-year shoot itself is also derived from an axillary bud, the first and second leaves correspond to prophylls, and the axillary buds of these prophylls become the shoot apices of the following year’s shoots. In contrast, the axillary buds of leaves other than the prophylls form aerial branches. The axillary bud of the first leaf becomes the innovation bud and elongates in the following year. The axillary bud of the second leaf becomes the reserve bud, which usually elongates when the innovation bud is damaged or dies. However, depending on environmental conditions and species, both the innovation bud and the reserve bud may elongate. Although the corms of Colchicum and Gloriosa differ greatly in overall appearance, they are homologous organs.
In Colchicum, the stem axis of the previous year’s shoot becomes swollen to form a corm. When the corm is dug up, it has an outer tunic derived from the leaf sheath of the first leaf and an inner tunic derived from the leaf sheath of the second leaf. The latter is often slightly paler than the former. When these tunics are removed at the end of summer, it can be seen that the innovation bud, which is the axillary bud of the first leaf, and the reserve bud, which is the axillary bud of the second leaf, have already begun to elongate.
In Gloriosa, as in Colchicum, the stem axis of the previous year’s shoot becomes swollen. However, unlike in Colchicum, the bases of the two axillary buds elongate into rod-like structures in Gloriosa. The axillary buds are located at the tips of these rod-like structures. Because each rod-like structure is formed by elongation of the stem axis of the previous year’s shoot at the base of an axillary bud, its surface bears no leaf traces and is smooth except for the apical region bearing the axillary bud. In the following spring, the innovation bud, which is the axillary bud of the first leaf, elongates to form a new shoot. Usually, only the bases of the axillary buds of the first and second leaves elongate, producing a bifurcate corm. Rarely, as shown in the photograph, the base of the axillary bud of the third leaf also elongates, in addition to those of the first and second leaves, producing a trifurcate corm.
Brantjes, N.B.M., and Bos, J.J. (1980). Hawkmoth behavior and flower adaptation reducing self pollination in two Liliiflorae. New Phytol. 84, 139–143. https://doi.org/10.1111/J.1469-8137.1980.TB00756.X;WGROUP:STRING:PUBLICATION.
Corbera, J., Alvarez-Cros, C., and Stefanescu, C. (2018). Evidence of butterfly wing pollination in the martagon lily Lilium martagon L. Butll. Inst. Catalana Hist. Nat. 82, 117–120.
Corbera, J., Alvarez-Cros, C., and Stefanescu, C. (2018). Evidence of butterfly wing pollination in the martagon lily Lilium martagon L. Butll. Inst. Catalana Hist. Nat. 82, 117–120.
Daniels, R.J., Johnson, S.D., and Peter, C.I. (2020). Flower orientation in Gloriosa superba (Colchicaceae) promotes cross-pollination via butterfly wings. Ann. Bot. 125, 1137–1149. https://doi.org/10.1093/AOB/MCAA048.
Nordenstam, B. (1998). Colchicaceae. In The Families and Genera of Vascular Plants. Vol. III. Monocotyledons Lilianae (except Orchidaceae)., K. Kubitzki, ed. (Springer), pp. 175–185.
Thi, N. P.A., Kim, J.S., and Kim, J.H. (2013). Molecular phylogenetic relationships and implications for the circumscription of Colchicaceae (Liliales). Botanical Journal of the Linnean Society 172, 255–269. https://doi.org/10.1111/BOJ.12037;WGROUP:STRING:PUBLICATION.
Weiss, M.R. (1997). Innate colour preferences and flexible colour learning in the
タヌキノショクダイ科 Thismiaceae タヌキノショクダイ Thismia abei: Cropped from a photo by Kenji Suetsugu, Suetsugu et al. (2026), CC BY 4.0; ヒナノボンボリ Oxygyne hyodoi: Cropped from a photo by Syozi Hyodo, Cheek et al. (2018) Fig. 6, CC BY 4.0
All species of Thismiaceae are mycoheterotrophic and non-photosynthetic, occurring on the dark forest floor from tropical to temperate regions, with their center of diversity in the tropics. The distribution maps in POWO are based on country-level records and therefore depict broader distributions than the actual species ranges.
コウベタヌキノショクダイの自生地 Natural habitat of Thismia kobensis ヒノキの植林にコナラなどの広葉樹が侵入した林内で、コウベタヌキノショクダイの生育地点周辺には他の草本植物はほとんど生育していない。6月に写真の人物がいる場所で昨秋の落葉を取り除くと、その下で開花している個体が観察された。The photographs above show a natural habitat of Thismia kobensis. The site is located in a Japanese cypress (Chamaecyparis obtusa) plantation that has been slightly invaded by broad-leaved trees such as Quercus serrata. Few other herbaceous plants occur in the vicinity of the T. kobensis population. When the leaf litter deposited during the previous autumn was removed at the location indicated by the person in the photographs in June, flowering individuals were found beneath it.
タヌキノショクダイ科に特有の形質 Distinctive characters of Thismiaceae
Thismiaceae is characterized by two shared characters: (1) all species are fully mycoheterotrophic, and (2) ectopic appendages are formed on at least one of the sepals, petals, or stamens (except in Haplothismia).
1. 完全菌従属栄養 Fully mycoheterotrophic Thismia macahensis, Thismia aseroe, Thismia (= Bognisia) episcopalis: Drawings are reproduced from Engler (1889); Thismia variabilis: Cropped from a photo by Mayk Oliveira, https://www.inaturalist.org/photos/595106823, CC BY-NC; Thismia hyalina: Cropped from a photo by cm_celeste, https://www.inaturalist.org/photos/468954877, CC BY-NC; Thismia hyalina:_Cropped from a photo by Martin Acosta, https://www.inaturalist.org/photos/126745926, CC BY-NC; Thismia huangii: Cropped from a photo by 羅元甫, https://www.inaturalist.org/photos/283029670, CC BY-NC; Thismia alba: Cropped from a photo by AtiwatBoonrit, https://www.inaturalist.org/photos/138703569, CC BY-NC; Thismia sp.: Cropped from a photo by Michelle Honey, https://www.inaturalist.org/photos/65865470, CC BY-NC; Relictithismia kimotsukiensis: Cropped from a photo from Suetsugu et al. (2024), CC BY 4.0
1. 完全菌従属栄養 Fully mycoheterotrophic
タヌキノショクダイ科Thismiaceaeの全種は完全菌従属栄養性 fully mycoheterotrophicである(塚谷 2016; 末次 2023)。多くの植物は複数種のアーバスキュラー菌根菌と共生し、光合成によって生産した炭水化物や脂質を菌類に供給する代わりに、菌類からリン、窒素、その他無機塩や水分を得ている(Parniske 2008)。一方、タヌキノショクダイ科の植物は光合成を行わないため、菌類に光合成産物を供給せず、生活に必要な炭素源を含む栄養分と水分を菌類から得ている(Gomes et al., 2017; Merckx et al., 2017)。菌類が利用する炭素源はもともと周辺の光合成植物に由来するため、タヌキノショクダイ科の植物は菌類を介して周辺の植物から間接的に炭素を獲得しているともいえる。共生するアーバスキュラー菌根菌は通常1種または少数種に限られ(Guo et al. 2019、Gomes et al. 2017)、周辺の光合成植物と比較して宿主特異性が高く、共生関係の特殊化が進んでいると考えられている(Gomes et al. 2017)。
タヌキノショクダイ科では、糸状根を出す塊根 tuberous roots with filiform roots、ミミズ形根 vermiform roots、サンゴ状根 coralloid rootsなどの根形態が知られている(Feller et al. 2022)。このような根形態の多様化は、(1) 菌糸が共生できる組織を増やすこと、(2) 広い表面積によって菌糸との接触、感染機会を高めること、(3) 菌由来の栄養分を短距離で効率よく植物体へ輸送すること、という三つの条件を最適化する方向に進化した結果であり、各タイプの根がそれぞれ異なる最適解に到達したものと考えられている(Feller et al. 2022) 。上図では、T. macahensis、T. hyalina、T. variabilisが塊根型、T. aseroe、T. alba、T. huangii,T. sp.がミミズ形根型、T. episcopalisおよびRelictithismiakimotsukiensisはサンゴ状根型である。
All species of Thismiaceae are fully mycoheterotrophic. Most plants form symbiotic associations with multiple species of arbuscular mycorrhizal fungi, supplying the fungi with carbohydrates and lipids produced by photosynthesis in exchange for phosphorus, nitrogen, other mineral nutrients, and water (Parniske 2008). In contrast, because members of Thismiaceae are non-photosynthetic, they do not provide photosynthates to their fungal partners but instead obtain their nutrients and water, including carbon sources required for growth and survival, from the fungi (Gomes et al. 2017; Merckx et al. 2017). Since these carbon sources ultimately originate from surrounding photosynthetic plants, members of Thismiaceae can be regarded as indirectly acquiring carbon from neighboring plants via their fungal associates. The arbuscular mycorrhizal fungi associated with Thismiaceae are usually restricted to one or a few species (Guo et al. 2019; Gomes et al. 2017), exhibiting greater host specificity than those associated with surrounding photosynthetic plants, and are therefore considered to represent highly specialized symbiotic relationships (Gomes et al. 2017).
Three major root morphologies are recognized in theThismiaceae: tuberous roots with filiform roots, vermiform roots, and coralloid roots (Feller et al. 2022). The diversification of these root types has been hypothesized to reflect optimization of three functional requirements: (1) increasing the amount of tissue available for fungal colonization, (2) enhancing opportunities for fungal contact and infection through a larger surface area, and (3) facilitating efficient transport of fungus-derived nutrients to the plant body over short distances. The different root types are therefore thought to represent alternative evolutionary solutions to these functional demands (Feller et al. 2022). In the figure above, T. macahensis, T. hyalina, and T. variabilis possess tuberous roots; T. aseroe, T. alba, T. huangii, and Thismia sp. possess vermiform roots; and T. episcopalis and Relictithismia kimotsukiensis possess coralloid roots.
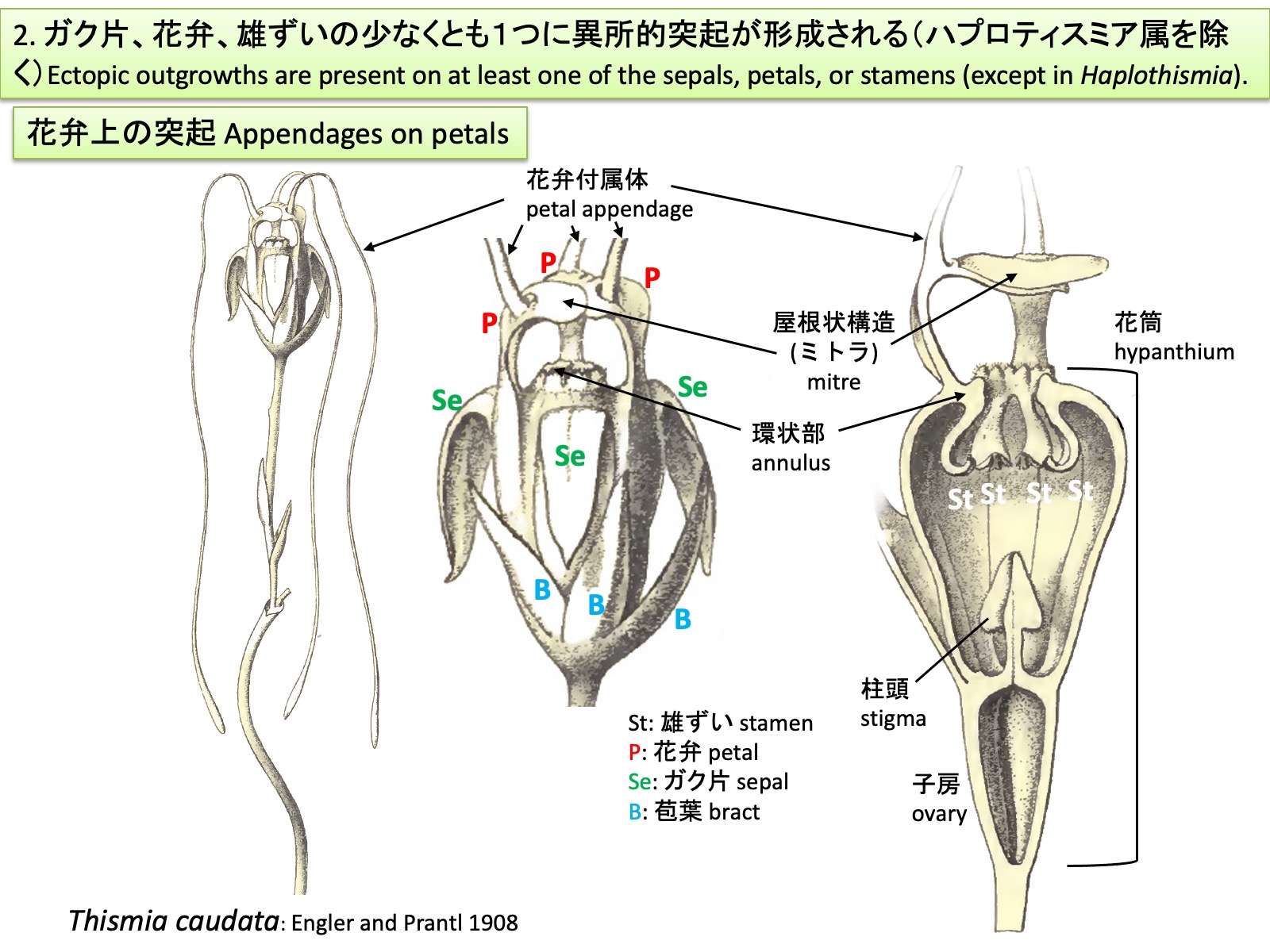
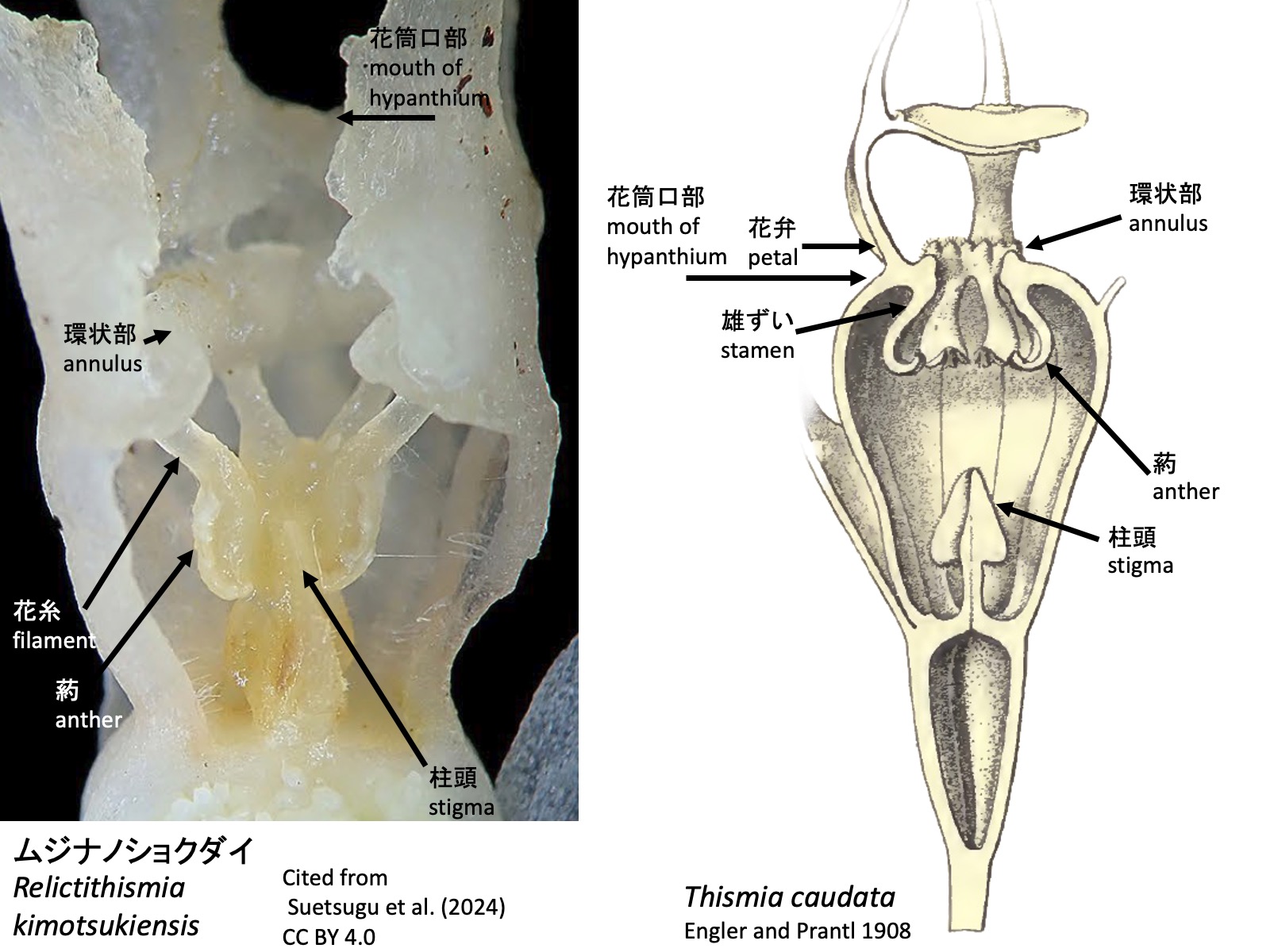
2. ガク片、花弁、雄ずいの少なくとも1つに異所的突起が形成される(ハプロティスミア属を除く) 2. Ectopic outgrowths are present on at least one of the sepals, petals, or stamens (except in Haplothismia) Thismia caudata: Drawings are reproduced from Engler and Prantl (1908)
2. ガク片、花弁、雄ずいの少なくとも1つに異所的突起が形成される(ハプロティスミア属を除く) Ectopic outgrowths are present on at least one of the sepals, petals, or stamens (except in Haplothismia).
タヌキノショクダイ科の花も他の被子植物の花と同様に、外側からガク片、花弁、雄ずい、雌ずいが形成される。ガク片は外花被、花弁は内花被とも呼ばれる。上図のブラジル産Thismia caudataでは、花弁から糸状の花弁付属体petal appendages が伸長するとともに、3枚の花弁の先端側が合着して屋根状構造 mitreを形成する(Engler and Prantl 1908; Ríos and da Cruz 2023)。Mitreはラテン語mitraに由来する英語表記で、日本語ではミトラと呼ばれることが多い。ガク片、花弁および雄ずいは癒合して花筒を形成し、その開口部は隆起して環状部annulusとなる。雄ずいは花筒内に下垂し、葯は花筒の外側に向かって裂開する。
Flowers of Thismiaceae, like those of other angiosperms, are composed of sepals, petals, stamens, and carpels arranged from the outside inward. The sepals are also referred to as the outer perianth, and the petals as the inner perianth. In the Brazilian species Thismia caudata, shown above, filamentous petal appendages elongate from the petals, while the distal portions of the three petals fuse to form a roof-like structure known as a mitre (Engler and Prantl 1908; Ríos and da Cruz 2023). The term “mitre” is derived from the Latin mitra and is often rendered in Japanese as mitora. The sepals, petals, and stamens are fused to form a floral tube, the opening of which is elevated into an annulus. The stamens hang down within the floral tube, and the anthers dehisce outward, away from the center of the floral tube.
ガク片上の突起 Appendages on sepals Thismia megalongensis: Cropped from a photo by izakschoon, https://www.inaturalist.org/photos/176122303, CC BY-NC
ガク片上の突起 Appendages on sepals
Thismia megalongensisでは、T. caudataと同様に、花弁付属体(水色矢印)と屋根状構造(黄緑色矢印)が形成される。さらに、ガク片からも突起(ガク片付属体:黄色矢印)が形成される。 In Thismia megalongensis, as in T. caudata, petal appendages (light blue arrows) and a mitre (light green arrows) are present. In addition, the sepals bear appendages (sepal appendages; yellow arrows).
雄ずい上の突起_1 Appendages on stamens_1 Thismia appendiculata: Drawings are reproduced from Schlechter (1919)
In many species of Thismia, the connective tissue and filaments of the stamens become elongated, resulting in fusion between adjacent stamens, expansion of their distal portions into laminar structures, or the formation of glands at the tips of the elongate structures. In Thismia appendiculata, shown above, the six stamens are fused to form a tube. In the upper right figure, a single stamen is highlighted in light blue. In the lower right figure, the anthers are shown in yellow.
雄ずい上の突起_2 Appendages on stamens_2 Thismia sp._1: Cropped from a photo by Michelle Honey, https://www.inaturalist.org/photos/65865480, CC BY-NC; Thismia sp._2: Cropped from a photo by Michelle Honey, https://www.inaturalist.org/photos/65865470, CC BY-NC
雄ずい上の突起_2 Appendages on stamens_2
タヌキノショクダイ属の種では、上図のThismia sp.のように、雄ずいの花糸基部が伸長して環状部 annulus を形成する。ムジナノショクダイ属Relictithismiaにおいても、環状部が認められるが(Suetsugu et al. 2024)、それがタヌキノショクダイ属と同様に雄ずいの花糸基部に由来するかどうかについては、発生過程がまだ報告されていない。
In species of Thismia, as in Thismia sp. shown above, the basal portions of the stamen filaments elongate to form the annulus. An annulus is also present in Relictithismia (Suetsugu et al. 2024), but its developmental process has not yet been reported, and it remains unclear whether it is derived from the basal portions of the stamen filaments, as in Thismia.
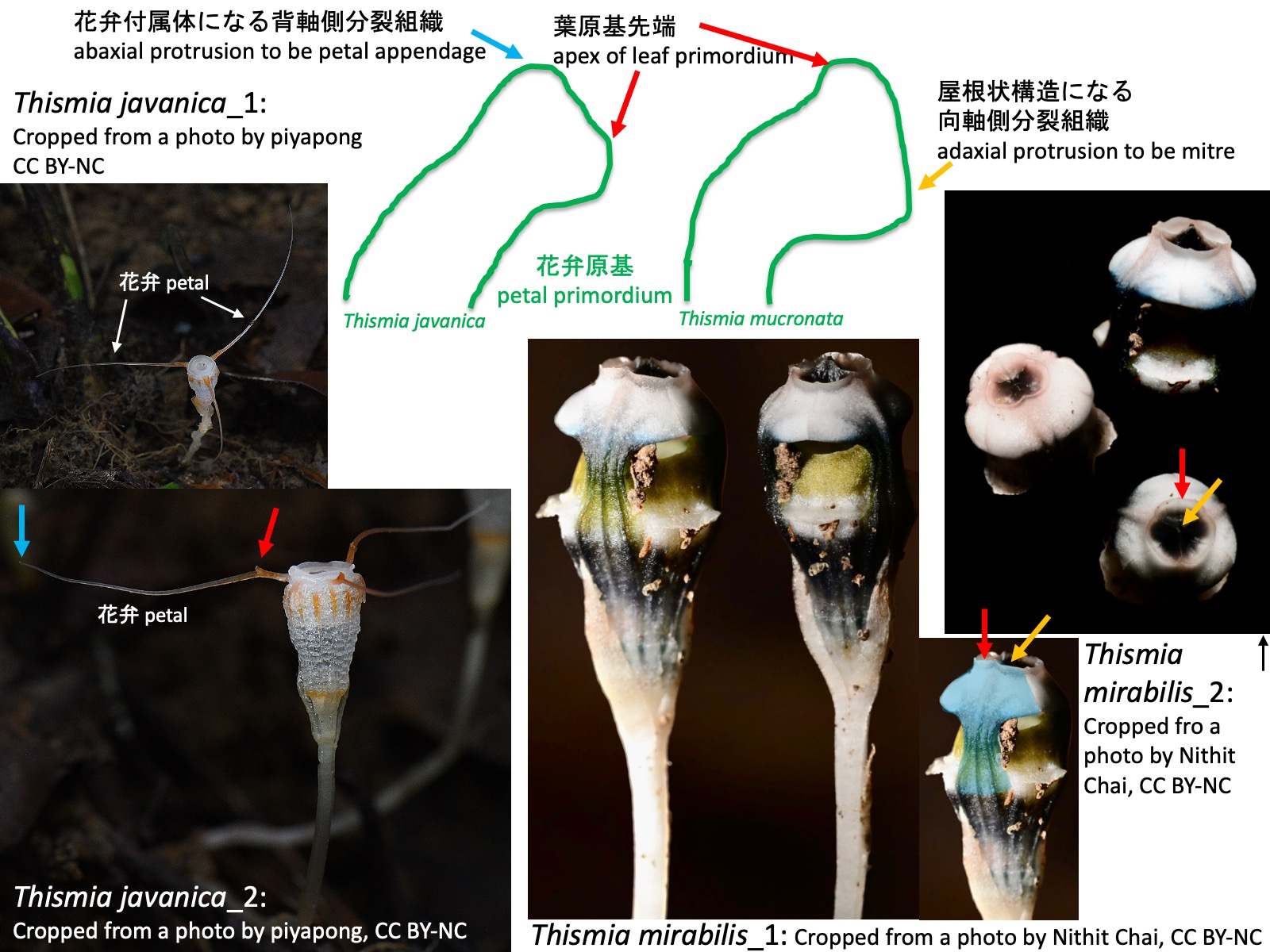
花弁付属体、屋根状構造、環状部の発生 Development of petal appendages, the mitre, and the annulus Thismia javanica_1: Cropped from a photo by piyapong, https://www.inaturalist.org/photos/441606101, CC BY-NC; Thismia javanica_2: Cropped from a photo by piyapong, https://www.inaturalist.org/photos/441609408, CC BY-NC; Thismia mirabilis_1: Cropped from a photo by Nithit Chai, https://www.inaturalist.org/photos/304715542, CC BY-NC; Thismia mirabilis_2: Cropped from a photo by Nithit Chai, https://www.inaturalist.org/photos/304715575, CC BY-NC
花弁付属体、屋根状構造、環状部の発生 Development of petal appendages, the mitre, and the annulus
ガク片、花弁、雄ずいの突起はどのように形成されるのだろうか。Nuraliev et al. (2021)は東南アジア産のThismia annamensis、T. javanica、T. mucronataの花発生過程を観察し、以下のことを報告した。
(1) T. javanicaでは、花弁原基の発生過程において、花弁原基の背軸側に新たな分裂組織が形成され、それが伸長して花弁付属体となる。上図左側にT. javanicaの花弁原基の模式図を示す。T. javanica_2の写真の青色矢印は花弁付属体の先端部を示している。一方、赤色矢印で示した突起についてはNuraliev et al. (2021)では言及されていない。この突起は花弁原基の先端部が伸長したものかもしれないが、その起源を明らかにするためには発生過程の観察が必要である。
(2) T. mucronataでは、花弁原基の発生過程において、花弁原基の向軸側に新たな分裂組織が形成され、それが伸長、癒合して屋根状構造となる。上図右側にT. mucronataの花弁原基の模式図を示す。Nuraliev et al. (2021)はT. mucronataを観察したが、上図にはShepeleva et al. (2020)においてT. mucronataの姉妹種とされ、類似した花形態をもつT. mirabilis の花の写真を示した。 右下の写真で青色に着色した部分は1枚の花弁に相当する。花弁先端部には隆起が認められ(赤色矢印)、さらにその先へ伸長している(黄色矢印)。赤色矢印で示した部分が花弁原基の先端部、黄色矢印で示した部分が向軸側に形成された分裂組織に由来する組織であるようにも見える。しかし、Nuraliev et al. (2021)では屋根状構造の形成過程そのものは観察されておらず、この点についてもさらなる発生学的研究が必要である。
How are the appendages of the sepals, petals, and stamens formed? Nuraliev et al. (2021) investigated floral development in the Southeast Asian species Thismia annamensis, T. javanica, and T. mucronata and reported the following observations.
(1) In T. javanica, during petal development, a new meristematic region is formed on the abaxial side of the petal primordium and subsequently elongates to form a petal appendage. A schematic diagram of the petal primordium of T. javanica is shown on the left side of the figure above. The blue arrow in photograph T. javanica_2 indicates the tip of a petal appendage. In contrast, the projection indicated by the red arrow was not mentioned by Nuraliev et al. (2021). This structure may represent an elongation of the distal portion of the petal primordium, but observations of its developmental process are required to clarify its origin.
(2) In T. mucronata, during petal development, a new meristematic region is formed on the adaxial side of the petal primordium. This tissue elongates and fuses to form a mitre. A schematic diagram of the petal primordium of T. mucronata is shown on the right side of the figure above. Although Nuraliev et al. (2021) studied T. mucronata, the figure presented here shows flowers of T. mirabilis, which was recovered as the sister species of T. mucronata by Shepeleva et al. (2020) and possesses a similar floral morphology. In the lower right photograph, the blue-colored region corresponds to a single petal. A prominent swelling is present near the petal apex (red arrow), and an elongate structure extends beyond it (yellow arrow). The region indicated by the red arrow appears to correspond to the distal portion of the petal primordium, whereas the structure indicated by the yellow arrow may be derived from the meristematic tissue formed on the adaxial side. However, the developmental stages leading to mitre formation were not observed by Nuraliev et al. (2021), and further developmental studies are needed to test this interpretation.
(3) Stamen primordia elongated toward the base of the hypanthium. The abaxial side of the basal portion of the staminal filaments elongated to form the annulus. In addition, ectopic elongation of the connective tissue and filaments gave rise to the complex morphology of the stamens. These observations suggest that at least some of the unusual floral structures found in Thismiaceae originated through the formation and subsequent elongation of ectopic meristematic regions on petal primordia, sepal primordia, staminal filaments, and connectives.
タヌキノショクダイ属 Thismia clade 5 :Thismia hexagona: Cropped from a photo by Vojtěch Tobias Blažek, https://www.inaturalist.org/photos/334762964, CC BY-NC; clade 4: Thismia mirabilis: Cropped from a photo by BioM_Akekachoke.B, https://www.inaturalist.org/photos/533284048, CC BY-NC; clade 2: Thismia rodwayi: Cropped from a photo by Katya Bandow, https://www.inaturalist.org/photos/166370615, CC BY-NC; clade 1: Thismia thaithongiana: Cropped from a photo by muangpaisuetrong, https://www.inaturalist.org/photos/317373844, CC BY-NC; Thismia panamensis: Cropped from a photo by lialavida, https://www.inaturalist.org/photos/424696457, CC BY-NC
タヌキノショクダイ属 Thismia
タヌキノショクダイ属について、ITS、18SrDNA、およびatpA遺伝子の塩基配列を用いた系統解析から、旧世界産の解析に用いられたタヌキノショクダイ属は単系統群であり、5つの単系統群(クレード)に分かれると推定されている(Shepeleva et al. 2020)。一方、新熱帯産のThismiapanamensisは別属の種Tiputiniafoetidaと姉妹群を形成することから、タヌキノショクダイ属全体は多系統群であると考えられている(Shepeleva et al. 2020)。
上図の系統樹は、18S rDNAおよびatpA遺伝子の塩基配列に基づいて推定されたものである(Suetsugu et al. 2024)。旧世界産タヌキノショクダイ属については、種名の代わりにクレード名を示し、各クレードに属する種の写真を併せて掲載した。
タヌキノショクダイ属の花形態は多様であり、いくつかの形質では収斂進化が生じている(Shepeleva et al. 2020)。そのため、形態形質に基づく属内分類系(Kumar et al. 2017)は、分子系統解析によって推定された系統関係(shepeleva et al. 2020, Suetsugu et al. 2024)と必ずしも一致しない。今後、より多くの種を対象とした分子系統解析が進むことで、系統関係を反映した属内分類体系が構築される可能性がある。以下では、タヌキノショクダイ属に見られる形態の多様性について概観する。
Phylogenetic analyses of ITS, 18S rDNA, and atpA sequences indicate that the sampled Old World species of Thismia form a monophyletic group that can be divided into five monophyletic clades (Shepeleva et al. 2020). In contrast, the Neotropical species Thismia panamensis forms a sister group to Tiputinia foetida, a species belonging to a different genus. Therefore, Thismia as currently circumscribed is considered to be polyphyletic (Shepeleva et al. 2020).
The phylogenetic tree shown above was inferred from 18S rDNA and atpA sequences (Suetsugu et al. 2024). For the Old World species of Thismia, clade names are shown instead of species names, and photographs of representative species belonging to each clade are presented alongside the tree. Floral morphology in Thismia is highly diverse, and several characters appear to have evolved convergently (Shepeleva et al. 2020). As a result, the infrageneric classification based on morphological characters (Kumar et al. 2017) does not necessarily correspond to the phylogenetic relationships inferred from molecular data (Shepeleva et al. 2020; Suetsugu et al. 2024). As molecular phylogenetic analyses become available for a greater number of species, it may become possible to establish an infrageneric classification that more accurately reflects evolutionary relationships. In the following sections, the morphological diversity of Thismia is reviewed.
タヌキノショクダイ属:ガク片付属体の有無 Thismia : Variation in sepal appendages Thismia taiwanensis: Cropped from a photo by 羅元甫, https://www.inaturalist.org/photos/411535969, CC BY-NC; Thismia huangii: Cropped from a photo by freesiahsu, https://www.inaturalist.org/photos/603575987, CC BY-NC
タヌキノショクダイ属:ガク片付属体の有無 Thismia : Variation in sepal appendages
タヌキノショクダイ属には、ガク片付属体を形成する種としない種がある。Thismiahuangii(左写真)ではガク片付属体は形成されないが、T. taiwanensis(右写真)ではガク片先端部が糸状に伸長し、ガク片付属体を形成する。花弁付属体と同様の発生過程を経るのであれば、この付属体はガク片先端部の背軸側に形成された分裂組織に由来する可能性がある。 Some species in Thismia possess sepal appendages, whereas others do not. In Thismia huangii (left photograph), sepal appendages are absent, whereas in T. taiwanensis (right photograph), the distal portion of the sepal elongates into a filamentous sepal appendage. If sepal appendages develop through a process similar to that of petal appendages, they may originate from a meristematic region formed on the abaxial side of the distal portion of the sepal.
タヌキノショクダイ属:屋根状構造の有無 Thismia : Variation in the mitre Thismia brunneomitroides: Cropped from a photo by piyapong, https://www.inaturalist.org/photos/441605563, CC BY-NC; Thismia aseroe: Cropped from a photo by Yee Chun Wah, https://www.inaturalist.org/photos/588094417, CC BY-NC
タヌキノショクダイ属:屋根状構造の有無 Thismia : Variation in the mitre
タヌキノショクダイ属には、屋根状構造を形成する種と形成しない種がある。Thismiabrunneomitroides (左写真)は屋根状構造を形成するのに対し、T. aseroe(右写真)では形成されない。 Within Thismia, some species form a mitre, whereas others do not. Thismia brunneomitroides (left photograph) possesses a mitre, whereas T. aseroe (right photograph) lacks this structure.
タヌキノショクダイ属:花弁付属体の有無_1 Thismia : Variation in petal appendages_1 Thismia arachnites: Cropped from a photo by กวินภพ ไชยยางพานิช, https://www.inaturalist.org/photos/446284372, CC BY-NC; Thismia selangorensis: Cropped from a photo by Joseph Pallante, https://www.inaturalist.org/photos/443856134, CC BY-NC
タヌキノショクダイ属:花弁付属体の有無_1 Thismia : Variation in petal appendages_1
タヌキノショクダイ属には、花弁付属体を形成する種と形成しない種がある。前図のThismiabrunneomitroidesでは花弁付属体は形成されないが、本図のThismia arachnites(左)およびThismia selangorensis(右)では花弁付属体が形成される。 Within Thismia, some species possess petal appendages, whereas others do not. In Thismia brunneomitroides shown in the previous figure, petal appendages are absent, whereas in Thismia arachnites (left) and Thismia selangorensis (right) shown in the present figure, petal appendages are present.
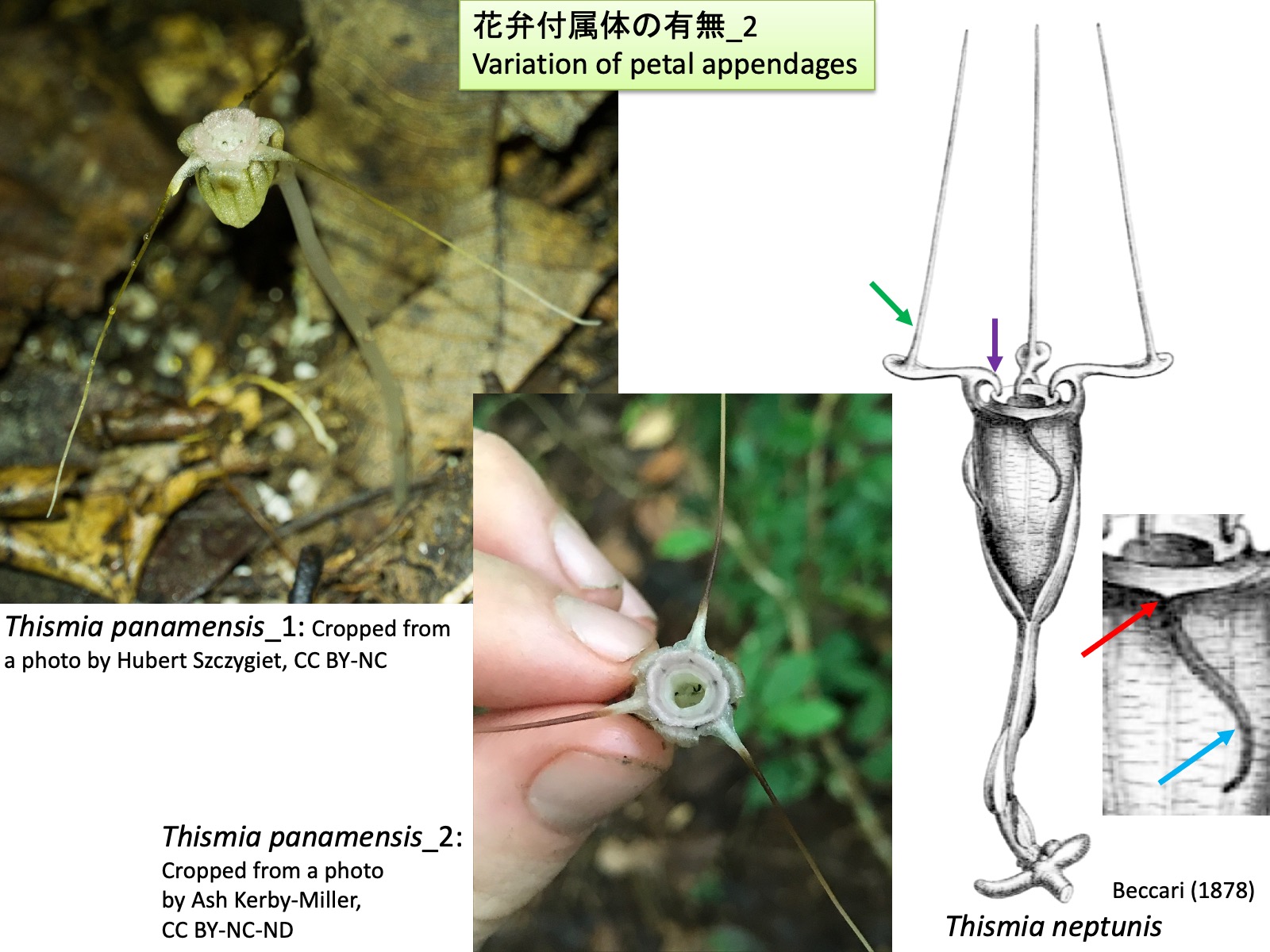
タヌキノショクダイ属:花弁付属体の有無_2 Thismia : Variation in petal appendages_2 Thismia panamensis_1: Cropped from a photo by Hubert Szczygiet, https://www.inaturalist.org/photos/81870711, CC BY-NC; Thismia panamensis_2: Cropped from a photo by Ash Kerby-Miller, https://www.inaturalist.org/photos/58695284, CC BY-NC-ND; Thismia neptunis: Drawings are reproduced from Beccari (1878)
タヌキノショクダイ属:花弁付属体の有無_2 Thismia : Variation in petal appendages_2
分子系統解析 (Shepeleva et al. 2020)において単系統群と推定された旧世界産タヌキノショクダイ属の種では、Nuraliev et al. (2021)が観察したように、花弁の背軸側または向軸側に新たな分裂組織が形成され、それが伸長して花弁付属体や屋根状構造を形成しているように見える。
さらに、旧世界であるボルネオ島産のThismia neptunisでは、前出のThismia megalongensisと同様に、ガク片付属体が形成される(Sochor et al. 2018)。もし花弁付属体と同様の発生過程を経るのであれば、ガク片向軸側基部付近に見られる突起 (赤色矢印)はガク片原基の先端部に相当し、長い突出部(水色矢印)はガク片原基の背軸側に形成された分裂組織に由来する可能性がある。花弁には、基部側の向軸側に鉤状突起 (紫色矢印)が形成される。この位置は、発生過程が観察された旧世界産タヌキノショクダイ属において屋根状構造を形成する突起(Nuraliev et al. 2021)の位置に類似しているが、形成される突起の形態は大きく異なる。さらに、花弁先端側の向軸側には細長い突起(緑色矢印)が形成される(Sochor et al. 2018)。これは、発生過程が解析された旧世界産タヌキノショクダイ属において、花弁付属体が花弁の背軸側に形成されること(Nuraliev et al. 2021)と対照的である。
In the Old World species of Thismia that were inferred to form a monophyletic group in molecular phylogenetic analyses (Shepeleva et al. 2020), petal appendages and mitres appear to be formed through the development of novel meristematic regions on either the abaxial or adaxial side of the petal primordia, followed by their elongation, as observed by Nuraliev et al. (2021).
In contrast, Thismia panamensis, a Neotropical species belonging to a lineage distinct from the Old World species of Thismia and forming a sister group to the genus Tiputinia, lacks the small projection at the adaxial base of the petal that is present in Old World species of Thismia and is thought to represent the original apex of the petal primordium. This observation suggests that the petal appendage of T. panamensis may not originate from an ectopic meristematic region formed on the abaxial side of the petal, but instead may represent an elongation of the original apex of the petal primordium itself. Petal appendages are absent in Tiputinia.
Furthermore, in Thismia neptunis from Borneo in the Old World, sepal appendages are formed (Sochor et al. 2018), as in the previously discussed T. megalongensis. If these structures develop through a process similar to that of petal appendages, the projection located near the adaxial base of the sepal (red arrow) may correspond to the apex of the sepal primordium, whereas the long appendage (light blue arrow) may be derived from a meristematic region formed on the abaxial side of the sepal primordium.
The petals bear hook-shaped projections on their adaxial side near the base (purple arrow). Although these projections occupy a position similar to that of the structures that give rise to the mitre in Old World species of Thismia whose development was studied by Nuraliev et al. (2021), the morphology of the resulting structures differs markedly. In addition, slender projections are formed on the adaxial side near the distal portions of the petals (green arrow) (Sochor et al. 2018). This contrasts with the Old World species of Thismia whose development was analyzed by Nuraliev et al. (2021), in which petal appendages are formed on the abaxial side of the petals. These observations suggest that both Old World and New World species of Thismia may include taxa in which floral appendages are formed through developmental pathways different from those documented in the Old World species studied by Nuraliev et al. (2021). Future developmental and phylogenetic studies will help to reassess infrageneric classification and to evaluate the homology of appendage structures among species.
タヌキノショクダイ属の送粉機構 Pollination mechanisms of Thismia Thismia tentaculata_1: Cropped from a photo by 桃子, https://www.inaturalist.org/photos/673952674, CC BY-NC; Thismia tentaculata_2: Cropped from a photo by 桃子, https://www.inaturalist.org/photos/673953422, CC BY-NC
タヌキノショクダイ属の送粉機構 Pollination mechanisms of Thismia
タヌキノショクダイ属の送粉昆虫についてはほとんど研究されていないが、Thismiatentaculataでは詳細な観察が行われている(Guo et al. 2019)。T. tentaculataの花には、キノコバエ類、ショウジョウバエ類、甲虫類、トビムシ類などの訪花が観察されたが、体に花粉を付着させており、有効な送粉者と推定されたのはクロバネキノコバエ科(Sciaridae)の Corynoptera 属のみであった(Guo et al. 2019)。また、日本に自生するタヌキノショクダイにおいても、クロバネキノコバエ科の種が訪花することが報告されている(Suetsugu et al. 2026)。
タヌキノショクダイ属の花の特異な色彩はキノコへの擬態である可能性が指摘されており、ガク片付属体、花弁付属体、屋根状構造は、キノコバエ類を誘引したり、花上を移動する際の足場として機能しているのではないかと考えられている(Guo et al. 2019)。
Although pollination biology of Thismia has attracted considerable interest, direct observations of pollinators are scarce because the plants are rare, small, and flower on the dark forest floor. Detailed observations have nevertheless been conducted for Thismia tentaculata (Guo et al. 2019). Visitors to the flowers of T. tentaculata included fungus gnats, drosophilid flies, beetles, and springtails. However, the only insects found carrying pollen and therefore considered effective pollinators belonged to the sciarid genus Corynoptera (Sciaridae) (Guo et al. 2019). Similarly, visits by sciarid flies have also been reported in the Japanese species Thismia kobensis (Suetsugu et al. 2026).
Pollinators are thought to be attracted by the yellow annulus of T. tentaculata. They enter the flower through the opening of the annulus (light blue arrows) and move through the narrow space between the stamens and the floral tube to reach the pistil located at the base of the floral tube. Because the anthers dehisce toward the floral tube, pollen is deposited on the body surface of the insect during this process. Droplets are secreted from hairs at the tips of the connectives, and these secretions have been suggested to function as rewards for pollinators. After reaching the base of the floral tube and contacting the sticky stigma, the insects eventually escape from the flower through the central tubular opening (*) surrounded by the stamens. The unusual coloration of Thismia flowers has been suggested to represent mushroom mimicry. The sepal appendages, petal appendages, and mitre may function in attracting fungus gnats and providing footholds for their movement on the flower (Guo et al. 2019).
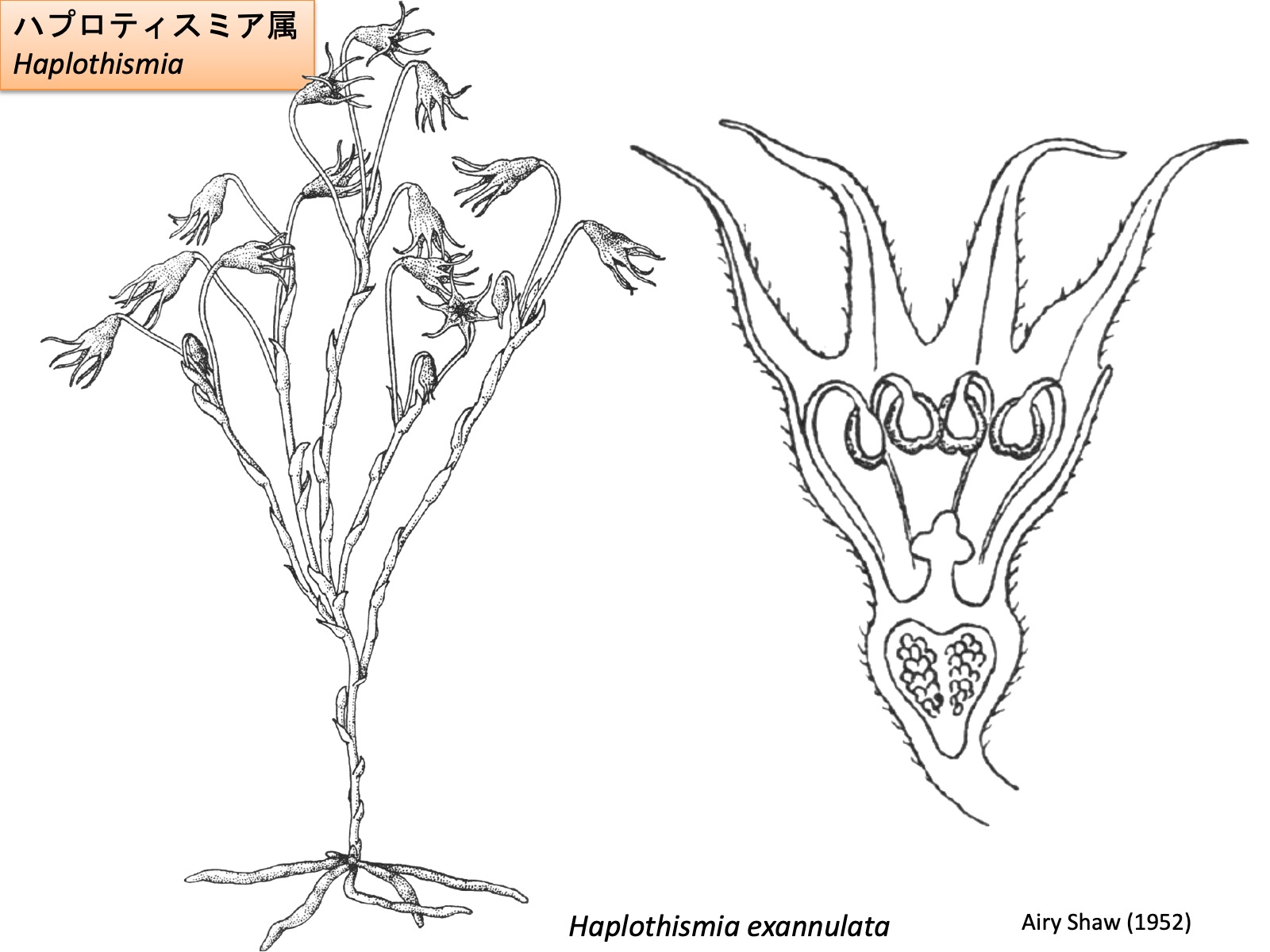
ハプロティスミア属 Haplothismia Drawings are reproduced from Airy Shaw (1952)
The flower of Haplothismia consists of three sepals, three petals, six stamens, and a single pistil composed of three fused carpels (Airy Shaw 1952). The sepals and petals are morphologically identical and can be distinguished only by their position. Unlike the other genera of Thismiaceae, Haplothismia does not produce ectopic appendages on its floral organs. The stamens are thought to elongate beyond the floral tube and subsequently bend inward toward its interior at their distal ends. In addition, the basal portions of the filaments do not undergo ectopic elongation, and no annulus is formed.
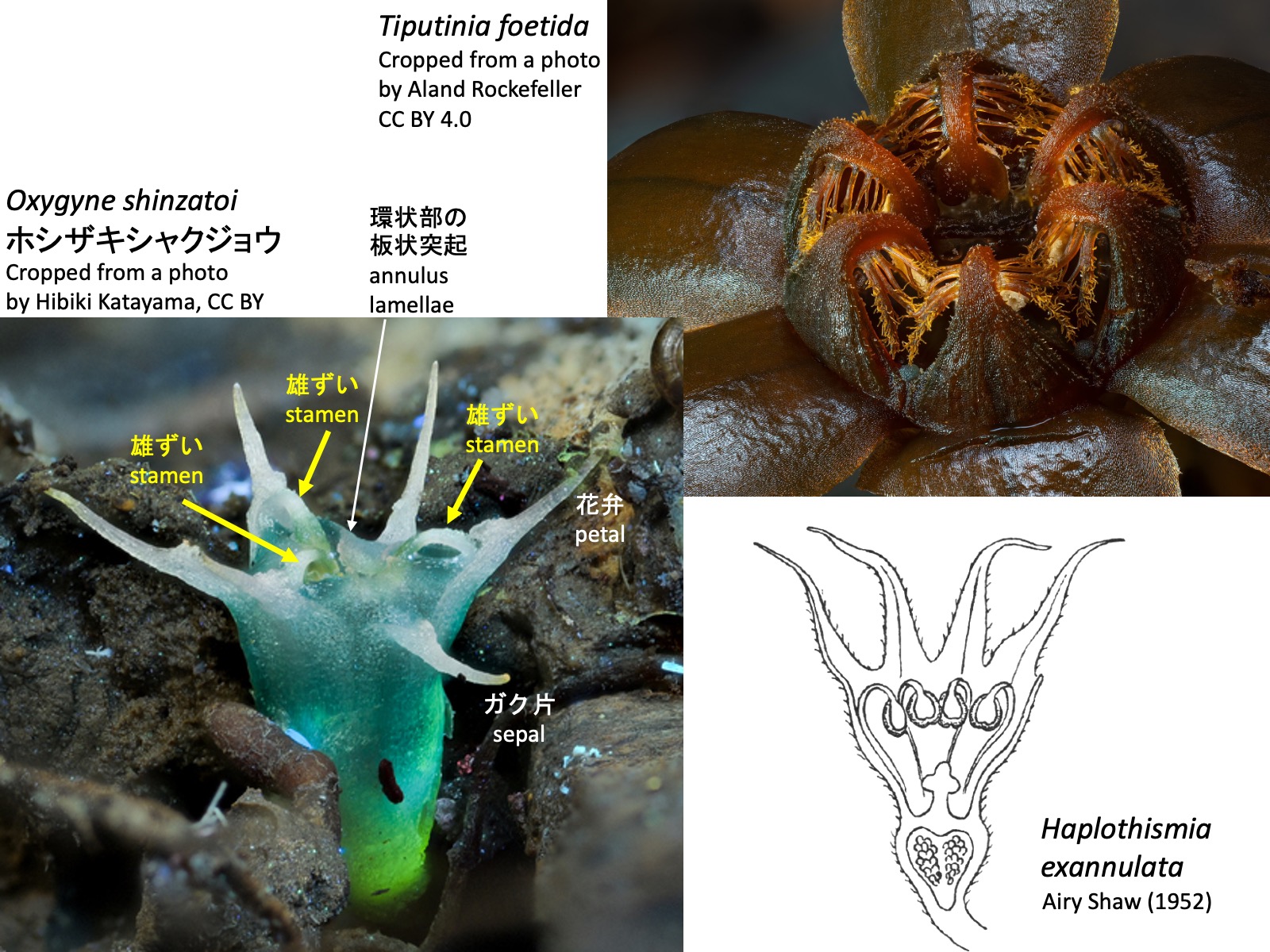
ヒナノボンボリ属 Oxygyne Oxygyne triandra: Drawings are reproduced from Engler, A. (1908); Oxygyne shinzatoi ホシザキシャクジョウ_1: Cropped from a photo by Hibiki Katayama, https://www.inaturalist.org/photos/328132754, CC BY; Oxygyne shinzatoi ホシザキシャクジョウ_2: Cropped from a photo by Hibiki Katayama, https://www.inaturalist.org/photos/328132800, CC BY; Oxygyne yamashitae ヤクノヒナホシ: Cropped from a photo by Kenji Suetsugu, Thorogood (2019), CC BY-NC-ND 4.0
ヒナノボンボリ属 Oxygyne
ヒナノボンボリ属の花も、3枚のガク片、3枚の花弁、3本の雄ずい、および3心皮が合着した1本の雌ずいからなる (Engler 1908)。ただし、環状部は板状突起によって形成される。Oxygyne triandraの右上図を見ると、この板状突起は花糸基部の向軸側が伸長して形成されたように見える。これは、タヌキノショクダイ属において花糸基部の背軸側が伸長して環状部の隆起を形成するのとは対照的である。さらに、雄ずいは花弁と対生する位置に形成されるが、雄ずいを欠くガク片対生位置にも板状突起が形成される (Yahara and Tsukaya 2008; Suetsugu et al. 2019)。もし板状突起が花糸基部に由来するのであれば、ガク片対生位置においても花糸基部の発生プログラムの一部が保持されている可能性がある。
The flowers of Oxygyne also consist of three sepals, three petals, three stamens, and a single pistil composed of three fused carpels. Unlike those of Thismia, however, the annulus is formed by annulus lamellae. As shown in the upper right figure of Oxygyne triandra, these annulus lamellae appear to be formed by elongation of the adaxial side of the filament base. This contrasts with Thismia, in which the abaxial side of the filament base expands to form the annular ridges. Furthermore, although stamens are produced opposite the petals, lamellar appendages are also present at positions opposite the sepals, where stamens are absent (Yahara and Tsukaya 2008; Suetsugu et al. 2019). If these lamellar appendages are indeed derived from the filament bases, this may indicate that part of the developmental program of the filament base is retained even at the sepal-opposed positions.
ムジナノショクダイ属 Relictithismia
ムジナノショクダイ属の花は、3枚のガク片、3枚の花弁、6本の雄ずい、および3心皮が癒合した1本の雌ずいからなる(Suetsugu et al. 2024)。ガク片と花弁は細長く伸長するが、タヌキノショクダイ属に見られるようなガク片上突起や花弁上突起は見られない。いずれも向軸側基部に機能不明な、丸い細胞からなるカルス状の隆起がある。この隆起は送粉昆虫の誘引に寄与しているのかもしれない。雄ずいは、花筒の口部で花弁から解離するのではなく、口部から全長の1/4ほど基部側の位置で解離する。環状部は雄ずいの解離部付近に形成される。右の写真(Suetsugu et al. 2024)を見ると、雄ずいはタヌキノショクダイ属と同様に、最初から下向きに伸長するように見える。
The flowers of Relictithismia consist of three sepals, three petals, six stamens, and a single pistil formed by the fusion of three carpels (Suetsugu et al. 2024). The sepals and petals are filiform, but they lack the sepal and petal appendages seen in Thismia. Both possess a callus-like cluster composed of rounded cells at the adaxial base, although its function remains unknown. This swelling may contribute to the attraction of pollinating insects. The stamens do not separate from the petals at the mouth of the floral tube, but instead separate at a position about one-quarter of the total length below the mouth. The annulus is formed at the region where the stamens separate. According to the right picture above (Suetsugu et al. 2024), the stamens appear to elongate downward from the outset, as in Thismia.
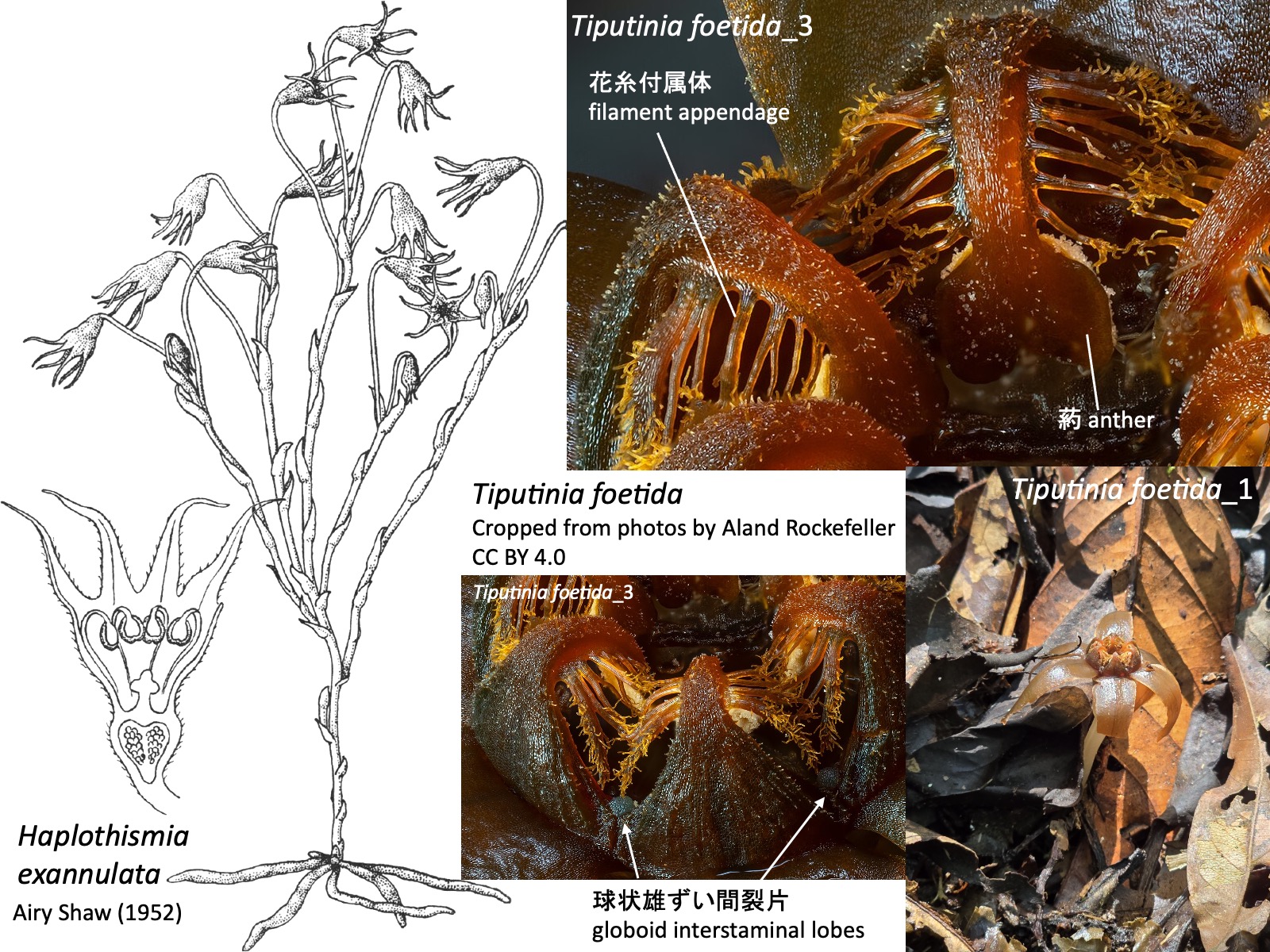
チプチニア属 Tiputinia Tiputinia foetida_1: Cropped from a photo by Alan Rockefeller, https://www.inaturalist.org/photos/362476435, CC BY; Tiputinia foetida_2: Cropped from a photo by Alan Rockefeller, https://www.inaturalist.org/photos/366763162, CC BY; Tiputinia foetida_3: Cropped from a photo by Alan Rockefeller, https://www.inaturalist.org/photos/367555871, CC BY
チプチニア属 Tiputinia
チプチニア属の花は、3枚のガク片、3枚の花弁、6本の雄ずい、および3心皮が癒合した1本の雌ずいからなる (Woodward et al. 2007)。雄ずいは花筒の外側へ伸び、途中で花筒の内側へ向けて屈曲する。花糸基部は広がって環状につながるが、花筒内側へ張り出す環状部annulusは形成されない。花糸からは花糸付属体が伸長し、雄ずい間には球状の雄ずい間裂片 globoid interstaminal lobesが形成される。
The flowers of Tiputinia consist of three sepals, three petals, six stamens, and a single pistil formed by the fusion of three carpels (Woodward et al. 2007). The stamens extend outward from the floral tube and then bend inward toward the inside of the tube. The filament bases are expanded and connected in a ring, but they do not form an annulus projecting into the floral tube. Filament appendages extend from the filaments, and globoid interstaminal lobes are formed between the stamens.
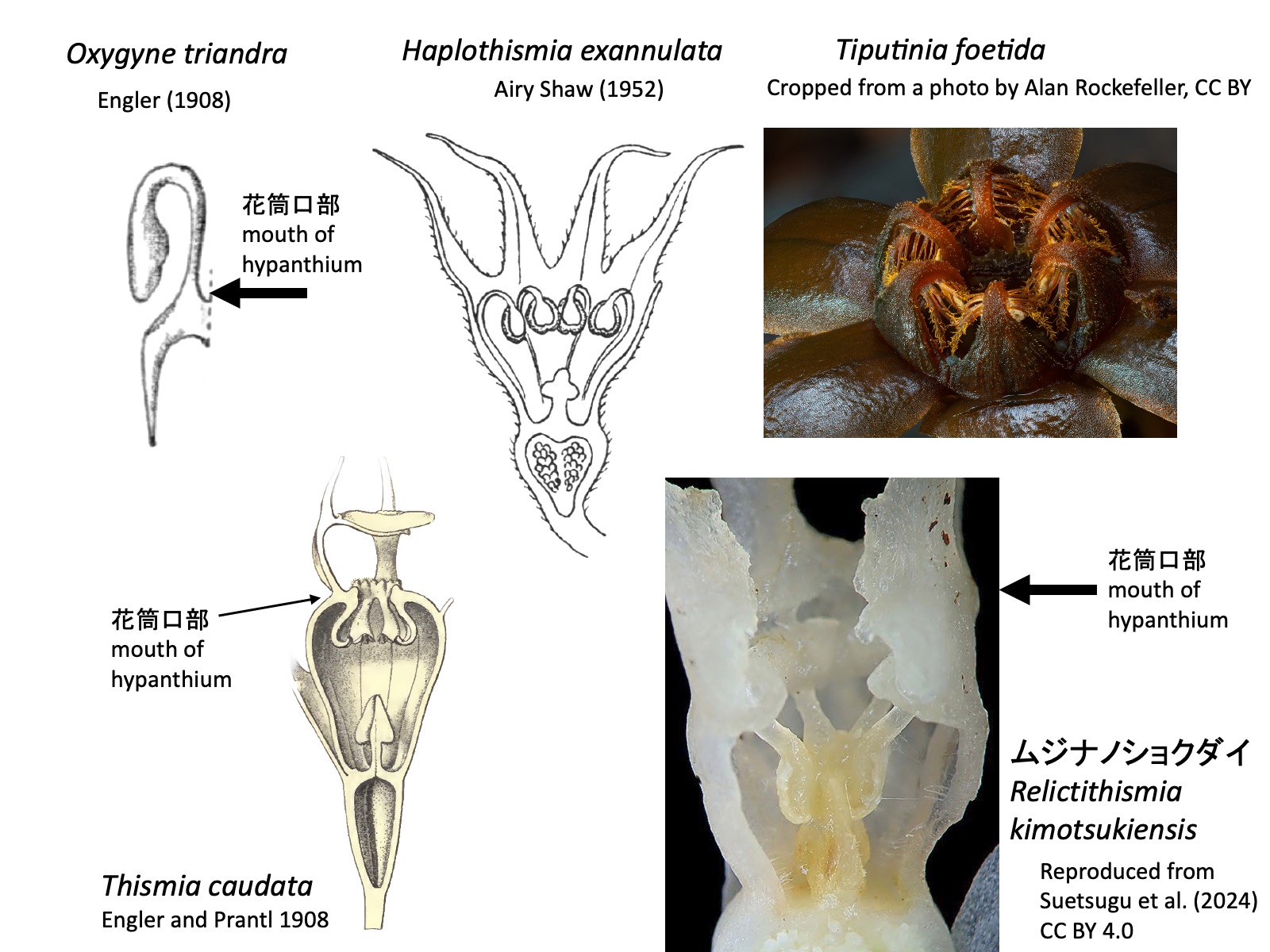
タヌキノショクダイ科の属の検索表(Cheek et al. 2018とSuetsugu et al. 2024を参考に作成) Key to the genera of Thismiaceae (based on Cheek et al. 2018 and Suetsugu et al. 2024)
1a. Staminal filaments free from the perianth at the mouth of the hypanthium. Stamens initially extend outward from the hypanthium but later bend inward; the bent portion is visible from outside the mouth of the hypanthium. Anthers positioned at the mouth of the hypanthium or slightly within it. …… 2 (Oxygyne, Haplothismia, Tiputinia)
1b. Staminal filaments free from the perianth within the hypanthium (in Relictithismia) or at the mouth of the hypanthium (in Thismia). Anthers positioned within the hypanthium. …… 4 (Relictithismia, Thismia)
Oxygyne triandra: Reproduced from Engler (1908); Haplothismia exannulata: Reproduced from Shaw (1952); Thismia caudata: Reproduced from Engler and Prantl (1908); Tiputinia foetida: Cropped from a photo by Alan Rockefeller, https://www.inaturalist.org/photos/367555871, CC BY;ムジナノショクダイ Relictithismia kimotsukiensis: Reproduced from Suetsugu et al. (2024) CC BY 4.0
Oxygyne shinzatoi ホシザキシャクジョウ: Cropped from a photo by Hibiki Katayama, https://www.inaturalist.org/photos/328132800, CC BY; Tiputinia foetida: Cropped from a photo by Aland Rockefeller, https://www.inaturalist.org/observations/205032669, CC BY 4.0; Haplothismia exannulata: Reproduced from Airy Shaw, H.K. (1952)
3a. Inflorescence aerial, repeatedly branched, with flowers borne at the tips of the branches. Stamens lacking ectopic appendages. …… Haplothismia
3b. Inflorescence produced close to the ground surface, usually bearing a single flower. Stamens with five or six pairs of filament appendages arising from the filaments. Globoid interstaminal lobes present. …… Tiputinia
Haplothismia exannulata: Reproduced from Airy Shaw (1952); Tiputinia foetida_1: Cropped from a photo by Alan Rockefeller, https://www.inaturalist.org/photos/362476435, CC BY; Tiputinia foetida_3: Cropped from a photo by Alan Rockefeller, https://www.inaturalist.org/photos/367555871, CC BY; Haplothismia exannulata: Reproduced from Airy Shaw (1952)
4a. Staminal filaments free from the perianth within the hypanthium, and the annulus formed within the hypanthium. The abaxial side of the anthers adheres to the stigma. …… Relictithismia
4b. Staminal filaments free from the perianth at the mouth of the hypanthium, and the annulus formed at the mouth of the hypanthium. Anthers separated from the stigma. …… Thismia
ムジナノショクダイRelictithismia kimotsukiensis: Reproduced from Suetsugu et al. (2024); Thismia caudata: Reproduced from Engler and Prantl (1908)
I am deeply grateful to Dr. Hirokazu Tsukaya, Dr. Kenji Suetsugu, Mr. Kazuma Takizawa, and Mr. Kazuki Inui for their invaluable help in preparing this account of Thismiaceae. Responsibility for all contents of this manuscript, however, rests solely with Mitsuyasu Hasebe.
引用文献 References
Airy Shaw, H.K. (1952). A new genus and species of Burmanniaceae from South India. Kew Bull. 7, 277. https://doi.org/10.2307/4109280.
Beccari, O. (1878) Burmanniaceae. Malesia 1: 240-253.
Cheek, M., Tsukaya, H., Rudall, P.J., and Suetsugu, K. (2018). Taxonomic monograph of Oxygyne (Thismiaceae), rare achlorophyllous mycoheterotrophs with strongly disjunct distribution. PeerJ : e4828. https://doi.org/10.7717/PEERJ.4828/FIG-10.
Engler, A. (1889) Burmanniaceae. In Engler, A. and Prantl, K. Die Natürlichen Pflanzen Familien. II Teil. 6. Abteilung. page 47, Fig. 38. Leipzig, Verlag von Wilhelm Engelmann.
Engler, A. (1908) Die Pflanzenwelt Afrikas insbesondere seiner tropischen Gebiete. Vol. 2 page 9. In: Engler & Drude (eds.), Die Vegetation der Erde 9. Leipzig: Wilhelm Engelmann.
Engler, A. and Prantl, K. 1908. Nat. Pflanzenfam. Erganzungshefte II. p. 73 Fig. 12. Thismia caudataがシノニムのGlaziocharis macahensisとして図示されている。Thismia caudata is illustrated under its synonym Glaziocharis macahensis.
Schlechter, R. (1919) 57. Eine neue Papuasische Burmanniacee. In Engler, A. (1919) Bot. Jahrb. Syst. 55: 202-203
Feller, B., Dančák, M., Hroneš, M., Sochor, M., Suetsugu, K., and Imhof, S. (2022). Mycorrhizal structures in mycoheterotrophic Thismia spp. (Thismiaceae): functional and evolutionary interpretations. Mycorrhiza 32, 269–280. https://doi.org/10.1007/S00572-022-01076-3.
Gomes, S.I.F., Aguirre-Gutiérrez, J., Bidartondo, M.I., and Merckx, V.S.F.T. (2017). Arbuscular mycorrhizal interactions of mycoheterotrophic Thismia are more specialized than in autotrophic plants. New Phytol. 213, 1418–1427. https://doi.org/10.1111/NPH.14249;WGROUP:STRING:PUBLICATION.
Guo, X., Zhao, Z., Mar, S.S., Zhang, D., and Saunders, R.M.K. (2019). A symbiotic balancing act: arbuscular mycorrhizal specificity and specialist fungus gnat pollination in the mycoheterotrophic genus Thismia (Thismiaceae). Ann. Bot. 124, 331–342. https://doi.org/10.1093/AOB/MCZ087.
Kumar, P., Gale, S.W., Li, J.-H., Bouamanivong, S., and Fischer, G.A. (2017). Thismia nigricoronata, a new species of Burmanniaceae (Thismieae, Dioscoreales) from Vang Vieng, Vientiane Province, Laos, and a key to subgeneric classification. Phytotaxa 319, 225–240. https://doi.org/10.11646/PHYTOTAXA.319.3.2.
Merckx, V.S.F.T., Gomes, S.I.F., Wapstra, M., Hunt, C., Steenbeeke, G., Mennes, C.B., Walsh, N., Smissen, R., Hsieh, T.H., Smets, E.F., et al. (2017). The biogeographical history of the interaction between mycoheterotrophic Thismia (Thismiaceae) plants and mycorrhizal Rhizophagus (Glomeraceae) fungi. J. Biogeogr. 44, 1869–1879. https://doi.org/10.1111/JBI.12994;PAGE:STRING:ARTICLE/CHAPTER.
Nuraliev, M.S., Yudina, S. V., Shepeleva, E.A., Truong, B.V., Do, T.X., Beer, A.S., and Remizowa, M. V. (2021). Floral structure in Thismia (Thismiaceae: Dioscoreales): new insights from anatomy, vasculature and development. Bot. J. Linn. Soc. 195, 501–531. https://doi.org/10.1093/BOTLINNEAN/BOAA066.
Parniske, M. (2008). Arbuscular mycorrhiza: the mother of plant root endosymbioses. Nat. Rev. Microbiol. 6, 10 6, 763–775. https://doi.org/10.1038/nrmicro1987.
Ríos, R.C., and da Cruz, V.J.M.V. (2023). Rediscovery of Thismia caudata after 129 years and synopsis of other achlorophyllous herbs from the Pico do Marumbi State Park and buffer zone in Piraquara, Paraná, Brazil. Rodriguésia 74, e01122022. https://doi.org/10.1590/2175-7860202374042.
Shepeleva, E.A., Schelkunov, M.I., Hroneš, M., Sochor, M., Dančák, M., Merckx, V.S., Kikuchi, I.A., Chantanaorrapint, S., Suetsugu, K., Tsukaya, H., et al. (2020). Phylogenetics of the mycoheterotrophic genus Thismia (Thismiaceae: Dioscoreales) with a focus on the Old World taxa: delineation of novel natural groups and insights into the evolution of morphological traits. Bot. J. Linn. Soc. 193: 287–315. https://doi.org/10.1093/BOTLINNEAN/BOAA017.
Sochor, M., Egertová, Z., Hroneš, M., and Dančák, M. (2018). Rediscovery of Thismia neptunis (Thismiaceae) after 151 years. Phytotaxa 340, 71–78. https://doi.org/10.11646/PHYTOTAXA.340.1.5.
末次健司(2023)「植物」をやめた植物たち(福音館書店)
Suetsugu, K., Sugimoto, T., and Tsukaya, H. (2019). Emended description and new localities of Oxygyne shinzatoi. (Burmanniaceae/Thismiaceae), with discussion of phylogenetic relationships of Oxygyne from Japan and Africa. Phytotaxa 423, 238-246. https://doi.org/10.11646/PHYTOTAXA.423.4.2.
Suetsugu, K., Nakamura, Y., Nakano, T., and Tagane, S. (2024). Relictithismia kimotsukiensis, a new genus and species of Thismiaceae from southern Japan with discussions on its phylogenetic relationship. J. Plant Res. 137: 411–422. https://doi.org/10.1007/S10265-024-01532-5.
Suetsugu, K., Sennikov, A.N., and Nuraliev, M.S. (2026). Multifaceted obscurity of Thismia abei (Thismiaceae): A fairy lantern with the protologue long disregarded in practice. Plants People Planet. https://doi.org/10.1002/PPP3.70214;JOURNAL:JOURNAL:25722611;REQUESTEDJOURNAL:JOURNAL:25722611;WGROUP:STRING:PUBLICATION.
Thorogood, C.J. (2019). Oxygyne: An extraordinarily elusive flower. Plants People Planet 1, 67–70. https://doi.org/10.1002/PPP3.26;JOURNAL:JOURNAL:25722611;REQUESTEDJOURNAL:JOURNAL:25722611;WGROUP:STRING:PUBLICATION.
塚谷裕一 (2016) 森を食べる植物 (岩波書店)
Woodward, C.L., Berry, P.E., Kamer, H.M. de, and Swing, K. (2007). Tiputinia foetida, a new mycoheterotrophic genus of Thismiaceae from Amazonian Ecuador, and a likely case of deceit pollination. Taxon 56, 157–162. https://doi.org/10.2307/25065746. Yahara, T., and Tsukaya, H. (2008). Oxygyne yamashitae, a new species of Thismiaceae from Yaku island, Japan. Acta Phytotax. Geobot. 59, 97–104. https://doi.org/10.18942/APG.KJ00005012322.
Fremontodendron californicum: photo by_garyashby, https://www.inaturalist.org/observations/652481152, CC BY-NC Chiranthodendron pentadactylon-1: photo by Erick Noe Tapia Banda, https://www.inaturalist.org/observations/17300607, CC BY-NC
フレモントデンドロン属 Fremontodendron とキラントデンドロン属 Chiranthodendron は姉妹群関係にあり、フレモントデンドロン連 Fremontodendreae を構成する(Baum et al. 2004)。この系統群は、アオイ亜科 Malvoideae およびパンヤ亜科 Bombacoideae と単系統群を形成するが、3者間の系統関係は十分には解明されていない(Baum et al. 2004; Colli-Silva et al. 2025)。フレモントデンドロン属の雄ずいは放射相称であるのに対し、キラントデンドロン属の雄ずいは左右相称で、手のような形態を示すため、学名のChiranthodendronは手のような花をつける木の意味で、デビルズハンドツリー“devil’s hand tree” と呼ばれています。
The genera Fremontodendron and Chiranthodendron are sister groups and together constitute the tribe Fremontodendreae (Baum et al. 2004). This clade forms a monophyletic group with the subfamilies Malvoideae and Bombacoideae, although the phylogenetic relationships among these three lineages remain unresolved (Baum et al. 2004; Colli-Silva et al. 2025). Whereas the stamens of Fremontodendron are radially symmetrical, those of Chiranthodendron are bilaterally symmetrical and resemble a hand, giving rise to the common name “devil’s hand tree.”
Hybrid between Fremontodendron and Chiranthodendron
フレモントデンドロン属Fremontodendron は3種からなり、アメリカ合衆国のカリフォルニア州およびアリゾナ州、ならびにメキシコのバハ・カリフォルニアに分布する(Kelman 1991)。デビルズハンドツリー と交雑し(Bayer and Kubitzki 2003)、写真のようにガク片が両属の中間的な色を呈し、左右相称の雄ずいを形成する。
Fremontodendron consists of three species distributed in California and Arizona in the United States and in Baja California, Mexico (Kelman 1991). It hybridizes with Chiranthodendron (Bayer and Kubitzki 2003), producing flowers in which the sepals show intermediate coloration between the two genera, as shown in the photograph, and the stamens become bilaterally symmetrical.
デビルズハンドツリーの自生地 The natural habitat of Chiranthodendron pentadactylon
キラントデンドロン属は、デビルズハンドツリーC. pentadactylon 1種からなり、メキシコ南部からグアテマラにかけての雲霧林などに点在して分布している(Hernández-Langford et al. 2020)。RAD-seq解析により、集団間の遺伝的分化が大きいことが示されており、かつてはより広範囲に分布していた種が、第四紀の気候変動を経て、各地に小規模な遺存集団として残存したと推定されている(Hernández-Langford et al. 2020)。写真は、メキシコ南部オアハカ州西部の標高約2550 mに位置する雲霧林の自生地。
The genus Chiranthodendron consists of a single species, C. pentadactylon, which is distributed discontinuously in cloud forests and related habitats from southern Mexico to Guatemala (Hernández-Langford et al. 2020). RAD-seq analyses revealed strong genetic differentiation among populations, suggesting that the species was formerly more widespread and that small relict populations persisted in scattered localities following Quaternary climatic changes (Hernández-Langford et al. 2020). The photograph shows a natural population in a cloud forest at an elevation of approximately 2550 m in western Oaxaca, southern Mexico.
民家の庭で栽培されているデビルズハンドツリー Chiranthodendron grown in a private garden
Possibly because it is used medicinally (Etkin 1994), it is sometimes planted in the gardens of private houses. The photograph shows an individual growing in the garden of a house located approximately 1 km from the natural population. Although this species can reach 40 m in height in the wild (Toledo 1975), this tree was approximately 15 m tall.
デビルズハンドツリーの花の構造 Flower structure of Chiranthodendron Chiranthodendron pentadactylon: Photo by Miguel García Cruz, https://www.inaturalist.org/photos/109313600, CC BY-NC
デビルズハンドツリーの花は、数枚の小苞葉に包まれ、厚く硬質な5枚のガク片を形成する(Bayer and Kubitzki 2003)。ガク片の外側(背側)は茶色の短毛で覆われ、内側は赤色で、基部には大きな黄色の蜜腺がある。5本の雄ずい(写真の*)は基部で癒合し、それぞれ細長く、先端に向かって細くなる。雄ずい筒は左右相称で、葯が花の下側に向かって開裂するように雄ずいがよじれている。雄ずい筒の中央から雌ずいが突出し、柱頭は葯の下側へ伸びる。
The flowers of Chiranthodendron pentadactylon are enclosed by several bracteoles and bear five thick, rigid sepals (Bayer and Kubitzki 2003). The outer (abaxial) surface of the sepals is covered with short brown hairs, whereas the inner surface is red and bears large yellow nectaries at the base. The five stamens are fused at the base, and each is elongated and tapers toward the apex. The staminal tube is bilaterally symmetrical, and the stamens are twisted so that the anthers dehisce toward the lower side of the flower. The pistil protrudes from the center of the staminal tube, and the stigma extends beneath the anthers.
デビルズハンドツリーの蕾の外観と内部の様子 External and internal structure of a Chiranthodendron flower bud
指のように伸びる雄ずいは、蕾の中ではげんこつ状に丸まっており、開花に伴って外側へ伸長する。
The finger-like stamens are curled into a fist-like shape within the flower bud and elongate outward as the flower opens.
The fruit, composed of a hard and thick ovary wall, becomes large enough to be barely grasped by an adult hand and contains numerous seeds inside.
デビルズハンドツリーの送粉者 Pollinators of Chiranthodendron Chiranthodendron pentadactylon-1: Photo by Sam Wilson, https://www.inaturalist.org/photos/251585038, CC BY-NC Chiranthodendron pentadactylon-2: Photo by Bernardino Villa B, https://www.inaturalist.org/photos/28370926, CC BY
Why does the devil’s hand tree possess such unusually shaped stamens? The presence of large nectaries and red, rigid sepals suggests that the flowers are bird-pollinated. Indeed, field observations in Guerrero, southern Mexico, have recorded visits by perching medium-sized birds such as orioles (Icterus) and New World warblers (Parulidae) (Toledo 1975). Hummingbirds have also been observed feeding from the flowers while perched rather than hovering (Toledo 1975). The five claw-like stamens are thought to function by guiding avian pollinators to enter the flower from a particular direction and by placing pollen on specific parts of the pollinator’s body, thereby increasing pollination efficiency (Toledo 1975). Because a space is present beneath the stamens, birds may enter from below and receive pollen on the head or dorsal side of the body; however, no direct observational report of this behavior could be found.
参考文献 References
Baum, D.A., Smith, S.D.W., Yen, A., Alverson, W.S., Nyffeler, R., Whitlock, B.A., and Oldham, R.L. (2004). Phylogenetic relationships of Malvatheca (Bombacoideae and Malvoideae; Malvaceae sensu lato) as inferred from plastid DNA sequences. Am. J. Bot. 91, 1863–1871. https://doi.org/10.3732/AJB.91.11.1863;WGROUP:STRING:PUBLICATION.
Bayer, C., and Kubitzki, K. (2003). Malvaceae. In The Families and Genera of Vascular Plants. (Volume eds.) K. Kubitzki and C. Bayer. Vol. V. Dicotyledons: Malvales, Capparales and Non-betalain Caryophyllales, K. Kubitzki, ed. (Springer), pp. 225–311.
Colli-Silva, M., Pérez-Escobar, O.A., Ferreira, C.D.M., Costa, M.T.R., Gerace, S., Coutinho, T.S., Yoshikawa, V.N., Antonio-Domingues, H., Hernández-Gutiérrez, R., Bovini, M.G., et al. (2025). Taxonomy in the light of incongruence: An updated classification of Malvales and Malvaceae based on phylogenomic data. Taxon 74, 361–385. https://doi.org/10.1002/TAX.13300;ISSUE:ISSUE:DOI.
Etkin, N.L. (1994). Eating on the Wild Side: The Pharmacologic, Ecologic and Social Implications of Using Noncultigens (Univ. Arizona Press).
Hernández-Langford, D.G., Siqueiros-Delgado, M.E., and Ruíz-Sánchez, E. (2020). Nuclear phylogeography of the temperate tree species Chiranthodendron pentadactylon (Malvaceae): Quaternary relicts in Mesoamerican cloud forests. BMC Evol. Biol. 20, 44-. https://doi.org/10.1186/S12862-020-01605-8/FIGURES/8. Kelman, W.M. (1991). A Revision of Fremontodendron (Sterculiaceae). Syst. Bot. 16, 3–20. https://doi.org/10.2307/2418969.
Lecythidaceae comprises approximately 30 genera and is widely distributed in the tropics (POWO 2026). It is characterized by the production of numerous stamens and includes genera in which part of the androecium is modified into staminodes, as well as genera in which the androecium has evolved into highly three-dimensional and diverse forms.
サガリバナ科の種は多数の雄ずいを持つ Species of Lecythidaceae have numerous stamens (Napoleonaea leonensis: Photo by Carel Jongkind, https://www.inaturalist.org/photos/79271253, CC BY-NC Scytopetalum pierreanum: Photo by Bart Wursten, https://www.inaturalist.org/photos/63908965, CC BY-NC Foetidia mauritiana: Photo by marc_roussin, https://www.inaturalist.org/photos/223220378, CC BY-NC)サガリバナ科の亜科の系統樹と分岐年代 Phylogeny and divergence times of Lecythidaceae subfamilies
サガリバナ科は、5亜科を含む広義のサガリバナ科として分類される場合(APG IV 2016;Stevens 2001 onwards)と、ナポレオナエア亜科およびスキトペタルム亜科をそれぞれ独立の科とし、残りの3亜科からなる狭義のサガリバナ科として分類される場合がある(Huang et al. 2015;Mori et al. 2017)。本稿では広義のサガリバナ科を採用する。系統樹の樹形および分岐年代は Rose et al. (2018) に基づく。
Lecythidaceae has been classified either in a broad sense, Lecythidaceae sensu lato including five subfamilies (APG IV 2016; Stevens 2001 onwards), or in a narrow sense, in which Napoleonaeoideae and Scytopetaloideae are treated as separate families and the remaining three subfamilies constitute Lecythidaceae sensu stricto (Huang et al. 2015; Mori et al. 2017). In this study, Lecythidaceae is treated in the broad sense. The topology and divergence times of the above phylogeny follow Rose et al. (2018).
2. Ovary superior or semi-inferior … Asteranthoideae
2. Ovary inferior or half-inferior … 3
3. Petals absent … Foetidioideae
3. Petals present … 4
4. Annular intrastaminal nectary present; distribution in the Old World … Barringtonioideae
4. Annular intrastaminal nectary absent; distribution in the New World … Lecythidoideae
ナポレオナエア亜科のナポレオナエア属とクラテランテゥス属 Napoleonaea and Crateranthus in Napoleonaeoideae Napoleonaea beninensis: Photo by Scott Zona, https://www.inaturalist.org/photos/91855160, CC BY-NC
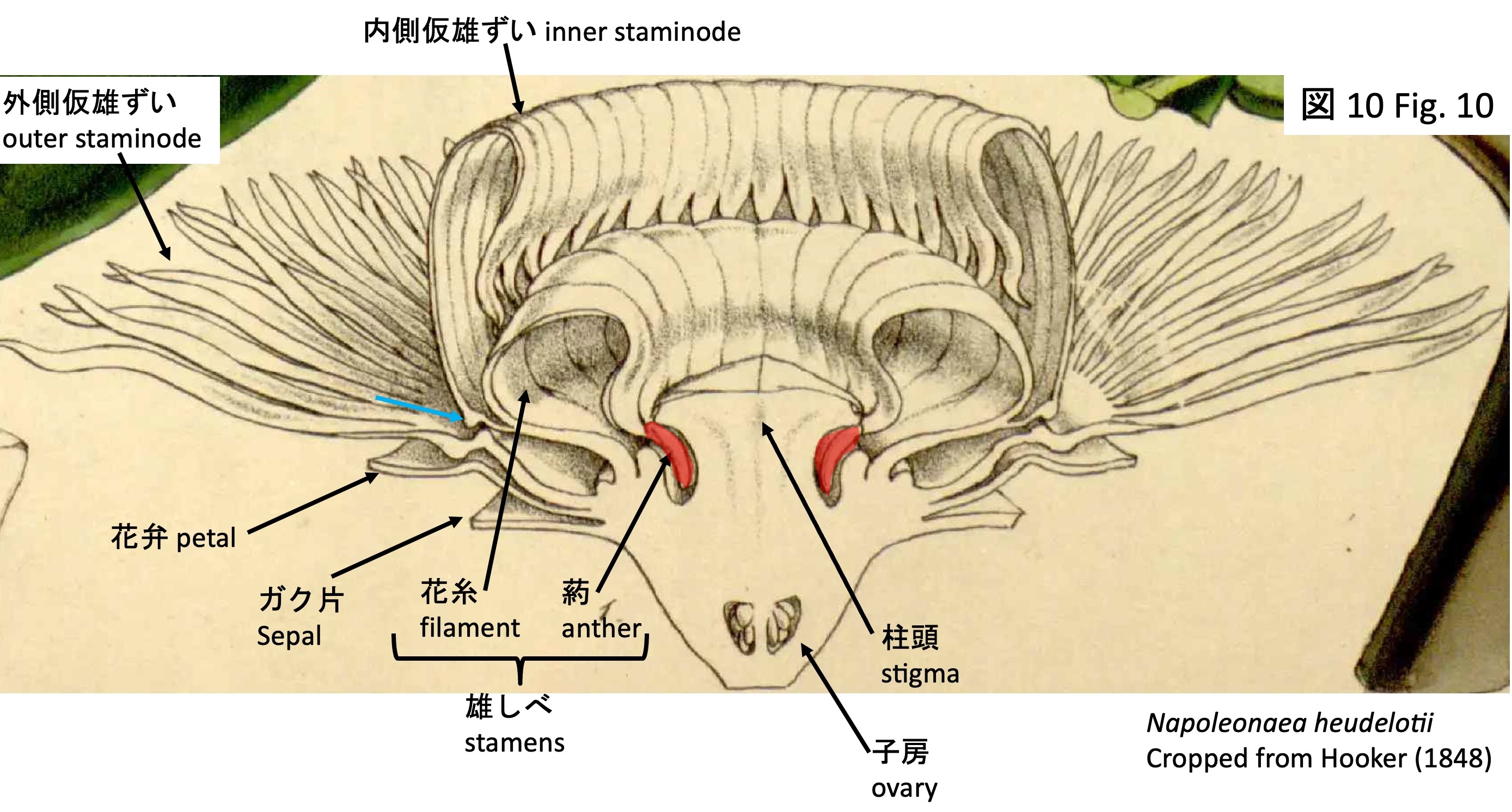
Napoleonaeoideae comprises two genera, Napoleonaea and Crateranthus (Mori et al. 2017). It is characterized by the production of numerous stamens, some of which are modified into petaloid staminodes. In the photographs, the petaloid staminodes are indicated by red arrows (Figs. 1, 2). Caption of the line drawing of Crateranthus latestui (Fig. 2) (Lecomte 1920): 1. Branch with leaves; 2. Bud; 3. Detached sepal; 4. Flower with sepals removed to show the staminodial corona (purple); 5. Same, viewed from above; 6. Longitudinal section of the flower before anthesis; 7. Longitudinal section of the flower after anthesis; 8. Transverse section of the ovary region; 9. Part of the staminodial corona (margin partly raised); 10. Stamens before and after anther dehiscence.
Both genera are distributed from West Africa to Central Africa.
Napoleonaea beninensisの花 A flower of Napoleonaea beninensis Napoleonaea beninensis: Photo by Scott Zona, https://commons.wikimedia.org/wiki/File:Napoleonaea_imperialis-2.jpg, CC BY 2.0.
ナポレオナエア属Napoleonaeaの花は、最も複雑な構造をもつ花の一つと考えられている(Frame and Durou 2001) (図5)。本属の特徴は、5裂した扁平な柱頭をもつことである(図6)。Napoleonaea beninensisの写真は記載の無いものは、名古屋市東山動植物園の栽培株を2026年4月22日に撮影させていただきました。心より感謝申し上げます。
The flowers of Napoleonaea are considered to be among the most structurally complex (Frame and Durou 2001) (Fig. 5). A distinctive feature of the genus is the presence of a broad, flattened, five-lobed stigma (Fig. 6). Unless otherwise noted, photographs of Napoleonaea beninensis were taken from a cultivated plant at Nagoya Higashiyama Zoo & Botanical Gardens on April 22, 2026. I sincerely thank them for their excellent cultivation.
Napoleonaea beninensisの花器官 Floral organs of Napoleonaea beninensis Napoleonaea heudelotiiの花の縦断面(Hooker 1848ではNapoleonaea imperialisと同定されていたが、Prance and Jongkind [2015]でN. heudelotiiに訂正された。) Longitudinal section of the flower of Napoleonaea heudelotii (identified as Napoleonaea imperialis in Hooker 1848, but later corrected to N. heudelotii by Prance and Jongkind [2015]).
Napoleonaea imperialisとN. beninensisの花の発生過程を調べた研究(Ronse de Craene 2011;原著でN. vogeliiとされているものは、Prance and Jongkind 2015によればN. beninensisに相当する)によると、花形成初期には数枚の小苞葉(図7)が形成され、その内側に生じる花原基から、まず5枚の萼片(図7)が輪状に形成される。次に、その内側に花弁(図8)が輪状に形成される。
花器官の原基形成順序は多様であり、典型的には萼片、花弁、雄ずい、雌ずいの順に形成されるが、雄ずいが花弁に先行する(萼片、雄ずい、花弁、雌ずい)場合も少なくない(Gifford and Foster 1989)。ところが、ナポレオナエア属では、花弁原基が伸長した後、雄ずいではなく花原基中央から雌ずい(図6)が形成される。その後、花弁と雌しべの間に雄ずい原基が輪状に伸長する。
Based on a study of floral development in Napoleonaea imperialis and N. beninensis (Ronse de Craene 2011; the material identified as N. vogelii in the original paper corresponds to N. beninensis according to Prance and Jongkind 2015), several bracteoles (Fig. 7) are first formed at an early stage of flower development. From the floral primordium arising inside these, five sepals (Fig. 7) are initiated in a whorl, followed by the whorled initiation of petals (Fig. 8).
The sequence of floral organ initiation is variable. Typically, organs arise in the order sepals, petals, stamens, and carpels, but in many cases stamens precede petals (sepals, stamens, petals, carpels). In Napoleonaea, however, after the petal primordia begin to elongate, the carpel primordia (Fig. 6) are initiated from the center of the floral primordium before the stamen primordia. Subsequently, stamen primordia develop in a whorl between the petal and carpel primordia.
The stamens (Fig. 9) are arranged as neighboring pairs of fertile stamens with developed anthers (F in Fig. 9) and sterile stamens lacking anthers (S in Fig. 9). Typically, both fertile and sterile stamens are positioned so as to lie beneath the flat, pentagonal stigma; in a sectional view of N. heudelotii (Fig. 10), the anthers inserted beneath the stigma are indicated in red. In the cultivated material examined here, however, the stigma is poorly developed, and the stamens are exposed (Fig. 9).
Subsequently, inner staminode primordia and then outer staminode primordia arise outside the stamen primordia.
No studies have reported the floral development of Crateranthus (Fig. 2). Petals are considered to be absent, with petaloid staminodes being formed instead (Lecomte 1920; Prance and Jongkind 2015). If these structures are homologous to the petaloid staminodes of Napoleonaea, they are likely to arise after the development of the carpel primordia, as in Napoleonaea. Conversely, if they are initiated earlier than the carpel primordia, they may be homologous to Napoleonaea petals.
However, the presence of dorsal projections (light blue arrows in Figs. 2 and 10), similar to those observed in the inner staminodes of Napoleonaea, suggests that these structures are most likely homologous. The function of these projections remains unknown in both genera.
Napoleonaea angolensisの送粉昆虫 Pollinators of Napoleonaea angolensis Napoleonaea angolensis-1: Photo by Bart Wursten, https://www.inaturalist.org/photos/63897726, CC BY-NC Napoleonaea angolensis-2: Photo by Anne-Hélène Paradis, https://www.inaturalist.org/photos/615702496, CC BY-NC
ナポレオナエア属の花が複雑化した理由として、送粉昆虫との共進化が考えられる。Napoleonaeaangolensisについては訪花昆虫が調べられている(図11、12)(Frame and Durou 2001; 注:Frame and Durou [2001]ではN. vogeliiとして記述されたが、Prance and Jongkind (2015)でN. angolensisに訂正された。)。
しかし、花室内にアザミウマ類を定着させる花は、Calluna(Hagerup 1950)やPeixotoa(Del-Claro et al. 1997)で知られているものの、ナポレオナエア属ほど複雑ではない。ナポレオナエア属の花がなぜこれほど複雑な構造をもつのかは、十分には説明されていない。
自生地では、観察した26個の花のうち、子房が膨らんだものは1個のみであり、結実率は低い。このことも踏まえると、過去にはアザミウマ類以外の送粉昆虫が存在し、それに適応していた可能性も考えられている(Frame and Durou 2001)。
The remarkable complexity of flowers in Napoleonaea is thought to reflect coevolution with pollinating insects. Insect visitors have been studied in N. angolensis (Figs. 11, 12) (Frame and Durou 2001; note: the material treated as N. vogelii in Frame and Durou [2001] was reassigned to N. angolensis by Prance and Jongkind [2015]).
At the bud stage, nectar is secreted from nectaries at the adaxial distal positions of the sepals (photographs of N. beninensis are shown in Figs. 13, 14), and ants are attracted to collect it. Attraction of ants is expected to provide some protection of young floral buds against herbivores; however, in natural populations about one-third of the buds are damaged by caterpillars, and the effectiveness of ant attraction remains unclear.
Flowers open around 7 p.m., but no insect visitors have been observed at that time. Around 8 a.m. the following morning, the flowers begin to emit a chocolate-like scent, and insect visitation increases. Thrips are attracted to aldehydes in the floral scent and to the white color; the space between the stigma and stamens (the floral chamber) provides a suitable refuge; and individuals bearing pollen have been observed within this chamber. These observations suggest that thrips are likely pollinators.
However, although flowers that harbor thrips within a floral chamber are known in Calluna (Hagerup 1950) and Peixotoa (Del-Claro et al. 1997), they are far less complex than those of Napoleonaea (Frame and Durou 2001). The reason for the extreme complexity of Napoleonaea flowers remains unclear.
In natural populations, only one out of 26 observed flowers showed ovary enlargement, indicating a low fruit set. Taken together, these observations raise the possibility that Napoleonaea may have originally been adapted to pollinators other than thrips (Frame and Durou 2001).
引用文献 References
Angiosperm Phylogeny Group (2016). An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG IV. Bot. J. Linn. Soc. 181, 1–20.
Del-Claro, K., Marullo, R., and Mound, L.A. (1997). A new Brazilian species of Heterothrips (Insecta: Thysanoptera) co-existing with ants in the flowers of Peixotoa tomentosa (Malphigiaceae). J. Nat. Hist. 31, 1307–1312.
Frame, D., and Durou, S. (2001). Morphology and biology of Napoleonaea vogelii (Lecythidaceae) flowers in relation to the natural history of insect visitors. Biotropica 33, 458–471. https://doi.org/10.1111/J.1744-7429.2001.TB00199.X;PAGE:STRING:ARTICLE/CHAPTER.
Gifford, E.M., and Foster, A.S. (1989). Morphology and Evolution of Vascular Plants 3rd ed. (W. H. Freeman and Company).
Hagerup, O. (1950). Thrips pollination in Calluna. Danske Biol. Meddel. 18, 1–16.
Huang, Y.Y., Mori, S.A., and Kelly, L.M. (2015). Toward a phylogenetic-based generic classification of neotropical Lecythidaceae – I. Status of Bertholletia, Corythophora, Eschweilera and Lecythis. Phytotaxa 203, 85-121–185. https://doi.org/10.11646/PHYTOTAXA.203.2.1.
Kandidov, M.V., Bobrov, A.V., Kandidov, M.V., and Bobrov, A.V. (2026). The history of distribution of the family Lecythidaceae s. l. Geogr. Environ. Sustain. https://doi.org/10.24057/2071-9388-2026-4560.
Lecomte, M.H. 1920. Deux especes nouvelles de genre Crateranthus (Lecythidacees). Bull. Muséum National d’Histoire Naturelle 26: 68-71.
Mori, S.A., Kiernan, E.A., Smith, N.P., Kelly, L.M., Huang, Y.-Y., Prance, G.T., and Thiers, B. (2017). Observations on the phytogeography of the Lecythidaceae clade (brazil nut family). Phytoneuron 30, 1–85.
Prance, G.T., and Jongkind, C.C.H. (2015). A revision of African Lecythidaceae. Kew Bull. 70, 1–68.
Ronse de Craene, L.P. (2011). Floral development of Napoleonaea (Lecythidaceae), a deceptively complex flower. In Flowers on the Tree of Life, L. Wanntorp and L. P. Ronse de Craene, eds. (Cambridge Univ. Press), pp. 279–295.
Rose, J.P., Kleist, T.J., Löfstrand, S.D., Drew, B.T., Schönenberger, J., and Sytsma, K.J. (2018). Phylogeny, historical biogeography, and diversification of angiosperm order Ericales suggest ancient Neotropical and East Asian connections. Mol. Phylogenet. Evol. 122, 59–79. https://doi.org/10.1016/J.YMPEV.2018.01.014.
Stevens, P. F. (2001 onwards). Angiosperm Phylogeny Website. Version 14. http://www.mobot.org/MOBOT/research/APweb/.
ツユクサ科の祖先的な花の花式図 Floral diagram of the ancestral condition in Commelinaceae
花式図は、図の上側が花序軸側、下側が苞葉側となるように描く (De Crane 2010)。ツユクサ科の花式図は上の左図に示すようになる (De Crane 2010, Evans et al. 2000)。右の写真はムラサキツユクサで花の構造を説明した。
Floral diagrams are drawn such that the upper side of the figure corresponds to the inflorescence axis and the lower side to the bract side (De Craene 2010). The floral diagram of Commelinaceae is as shown in the left panel (De Craene 2010; Evans et al. 2000). The photograph on the right shows Tradescantiaohiensis, in which the floral structure is illustrated.
Cochliostema odoratissimumは、中米南部から南米北部に分布し、タンク型(葉腋に水を溜める)着生植物で、直径5 cmに達するツユクサ科最大の花を付ける(Hardy and Stevenson 2000)。
Cochliostemaodoratissimum is distributed from southern Central America to northern South America. It is a tank-forming epiphytic plant (retaining water in the leaf axils) and produces the largest flowers in Commelinaceae, reaching up to 5 cm in diameter (Hardy and Stevenson 2000).
Cochliostema odoratissimum-1: Photo by Kabir Montesinos, https://www.inaturalist.org/photos/596297084, CC BY-NC-ND 4.0
Cochliostema odoratissimum-2: Photo by Diego cabrera, https://www.inaturalist.org/photos/466361117, CC BY-NC
Cultivated plants at the Higashiyama Zoo and Botanical Gardens, Nagoya, were used for the observations. We thank the Garden for granting permission to conduct the observations and for assistance with the tuning-fork experiments.
Cochliostema odoratissimumの花とツユクサ科の祖先的な花の比較 Comparison between the flower of Cochliostema odoratissimum and the ancestral flower of Commelinaceae
Cochliostema odoratissimumの花(右上の花式図と下方の写真)は、ツユクサ科の祖先的な花(左上の花式図)と比べて、1)1本の外側雄ずいと2本の内側雄ずい、計3本の雄ずいが癒合して、フード状雄ずい束(staminal cuculli:Hardy and Stevenson 2000)を形成すること、2)外側雄ずいのうち花序軸側の2本が長毛で覆われた外側仮雄ずいとなること、3) 花序軸側の内側雄ずいの原基はできるが成長しないこと、の3点で大きく異なる (Hardy and Stevenson 2000)。また、フード状雄ずい束の外側雄ずいの基部に橙色の長毛が密生する。さらに、花弁の縁に多細胞の数珠状毛が密生する形質は(右下写真)、コクリオステマ亜連 Cochliostematinaeの共有派生形質である (Hardy and Faden 2004)。
The flowers of Cochliostema odoratissimum (floral diagram at upper right and photographs below) differ markedly from the ancestral flowers of Commelinaceae (floral diagram at upper left) in three respects: (1) one outer stamen and two inner stamens (three in total) are fused to form a hood-like staminal fascicle (staminal cuculli; Hardy and Stevenson 2000); (2) of the outer stamens, the two on the inflorescence-axis side become outer staminodes covered with long hairs; and (3) the primordium of the inner stamen on the inflorescence-axis side is formed but does not develop (Hardy and Stevenson 2000). In addition, dense orange long hairs occur at the base of the outer stamens of the hood-like staminal fascicle. Furthermore, the presence of dense multicellular moniliform hairs along the petal margins (lower right photograph) is a synapomorphic character of Cochliostematinae (Hardy and Faden 2004).
The flowers of Cochliostema odoratissimum, as shown in the left figure, are formed with the staminal cuculli positioned more toward the basal side of the inflorescence axis than the pistil. If the inflorescence were to elongate linearly in this configuration, the staminal cuculli would be positioned on the lower side of the flower and the pistil on the upper side. In reality, however, the inflorescence axis twists in the scorpioid cyme, so that at anthesis the staminal cuculli are positioned on the upper side of the flower and the pistil on the lower side, as shown in the middle figure and the right photographs. When the inflorescence extends laterally (lower right photograph), as well as when it extends upward (upper right photograph), the pistil is consistently positioned on the lower side and the staminal cuculli on the upper side. This suggests that the pedicel may possess a mechanism for sensing gravity and controlling orientation.
Cochliostema odoratissimumのフード状雄ずい束の構造 Structure of staminal cuculli
フード状雄ずい束は、花の正面から見て、左側にねじれている(左上写真)。フード状雄ずい束を雌しべ側(左上写真の青色矢印、図B1から図B3)とその反対側(同緑色矢印、図G1から図G4)から観察した。(B1)雌しべ側では、2本の内側雄ずいが密に接しているが、組織としては合着していない。ピンセットを差し込むことで、こじ開けることができる。ただし、長毛が密生しており、フード状雄ずい束の内外で空気の移動はおこりうるものの、内側から外側へ花粉が放出されることは困難であるように見える。(G1、G2)雌しべから遠い側では、外側雄ずいの花糸と2本の内側雄ずいの花糸(G2の黒色星印)が癒合しており、G2の橙色点線部分に切れ目を入れなければ雄ずいを分離できない。分離した3本の雄ずいを、雌しべ側(B2)および反対側(G3、G4)から観察すると、フード状雄ずい束の内部は螺旋状の葯と白色の粉状の花粉(B3の黒色矢印)で満たされている。Cochliostemaは「螺旋状の雄ずい」を意味し、発生過程においてで葯の先端側が伸長し、4〜5回螺旋状に巻く(Hardy and Stevenson 2000、S1、S2)。S2の橙色矢印は、フード状雄ずい束内側に伸びだした花糸を示す。葯は全長にわたって縦裂開する。橙色の毛は外側雄ずいの基部から生じる。
The staminal cuculli are twisted to the left when viewed from the front of the flower (upper left photograph). The staminal cuculli were observed from the pistil side (blue arrow in the upper left photograph; Figs. B1–B3) and from the opposite side (green arrow; Figs. G1–G4). (B1) On the pistil side, the two inner stamens are closely appressed but are not fused as tissues. They can be pried apart by inserting forceps. However, although dense long hairs are present and air movement may occur between the inside and outside of the staminal cuculli, it appears difficult for pollen to be released from the inner to the outer side. (G1, G2) On the side distant from the pistil, the filament of the outer stamen is fused with the filaments of the two inner stamens (black asterisk in G2), and the stamens cannot be separated without making an incision along the orange dotted line in G2. When the three separated stamens are observed from the pistil side (B2) and from the opposite side (G3, G4), the interior of the staminal cuculli is filled with spiral anthers and white, powdery pollen (black arrow in B3). Cochliostema means “spiral stamens,” and during development the distal end of the anther elongates and coils spirally four to five times (Hardy and Stevenson 2000; S1, S2). The orange arrow in S2 indicates a filament extending into the interior of the staminal cuculli. The anthers dehisce longitudinally along their entire length. The orange hairs arise from the base of the outer stamens.
Cochliostema odoratissimumの送粉昆虫や送粉様式は報告されていないが(Hardy and Stevenson 2000)、開花するだけでは花粉がフード状雄ずい束の外側に放出されないことから、ハチ類がフード状雄ずい束に振動を与えることで花粉が放出される振動送粉(buzz-pollination)である可能性が指摘されている(Vogel 1978, Endress 1994, p. 155)。振動送粉は、無駄な花粉損失を防ぎ、適切な振動を生じさせることができる送粉者を選択できる利点があり(Mesquita-Neto aet al. 2018)、少なくとも2万種の被子植物で見られる(Vallejo-Marin and Russell 2024, Woodrow et al. 2024)。これらの花は、孔開葯 poricidal antherと呼ばれる、細長く、全体が裂開するのではなく先端の孔から花粉が徐々に放出される葯を持つことがほとんどである(Vallejo-Marin and Russell 2024, Woodrow et al. 2024)。
振動送粉をするハチ類は、葯に到達すると体をC字状に曲げて葯の周囲に巻き付き、胸部にある間接飛翔筋 indirect flight muscleの動きによって胸部、脚、あるいは頭部に振動を生じさせ、葯から花粉を放出させる(Vallejo-Marin and Russell 2024, Woodrow et al. 2024)。
Pollinating insects and the pollination system of Cochliostemaodoratissimum have not been reported (Hardy and Stevenson 2000). However, because pollen is not released to the outside of the staminal cuculli simply upon anthesis, it has been suggested that this species may be buzz-pollinated, with bees inducing pollen release by applying vibrations to the staminal cuculli (Vogel 1978; Endress 1994, p. 155). Buzz pollination has the advantages of preventing wasteful pollen loss and enabling the selection of pollinators capable of generating appropriate vibrations (Mesquita-Neto et al. 2018), and it occurs in at least 20,000 angiosperm species (Vallejo-Marín and Russell 2024; Woodrow et al. 2024). In most such flowers, the anthers are poricidal (poricidal anthers), being elongate and releasing pollen gradually through apical pores rather than by dehiscing along their entire length (Vallejo-Marín and Russell 2024; Woodrow et al. 2024).
In contrast, C. odoratissimum differs in that three stamens are fused to form a bivalve-like structure. The anthers are not poricidal but dehisce longitudinally along their entire length. In typical buzz-pollinated flowers, the yellow anthers themselves attract bees, whereas in C. odoratissimum the anthers are enclosed within the staminal cuculli and are not visible externally. Instead, orange hairs resembling anthers are present at the base of the outer stamens (orange arrows in the black-and-white and upper right photographs), and these are thought to attract bees (Vogel 1978). As indicated by the specific epithet, the petals emit a fragrance, although its role in attraction is unknown. Nectar is not secreted. The upper left photograph shows a frontal view and the lower photograph an oblique view; because the staminal cuculli are curved to the left when viewed from the front, the orange hairs are visible at the back right of the flower.
In buzz pollination, bees reaching the anthers curl their bodies into a C-shape around them and, through the action of the indirect flight muscles in the thorax, generate vibrations of the thorax, legs, or head that cause pollen to be released from the anthers (Vallejo-Marín and Russell 2024; Woodrow et al. 2024).
Even when the staminal cuculli of Cochliostema odoratissimum are shaken with forceps, little pollen is released.
ハチ類の振動は100から400 Hz程度であることから(De Luca et al. 2019, De Luca et al. 2012)、256 Hzの音叉を用いてフード状雄ずい束に振動を与えた。
Because bee vibrations are on the order of 100–400 Hz (De Luca et al. 2012, 2019), we applied vibrations to the staminal cuculli using a 256 Hz tuning fork.
https://youtu.be/tXUffPlYOgE
https://youtu.be/hlZY_BfEcuo
256 Hzの音叉によるCochliostema odoratissimumのフード状雄ずい束からの花粉放出(2回目) Pollen release from the staminal cuculli of Cochliostema odoratissimum induced by a 256 Hz tuning fork (second trial)
When the tuning fork was brought into contact with the pistil-side surface of the staminal cuculli, pollen was released vigorously. Approximately 10 seconds later, when the same staminal cuculli were stimulated again with the tuning fork, additional pollen was released, although in a smaller amount than in the first trial. No pollen release was observed after the third and subsequent stimulations.
https://youtu.be/3yK15U1nRE0
256 Hzの音叉によるCochliostema odoratissimumの別のフード状雄ずい束からの花粉放出 Pollen release from another staminal cucullus of Cochliostema odoratissimum induced by a 256 Hz tuning fork
In another flower as well, pollen release was observed when vibrations were applied with a tuning fork. In typical buzz pollination, pollen is released gradually from apical pores of the anthers, whereas in Cochliostema odoratissimum pollen is ejected forcefully from the staminal cuculli. What is the source of this driving force?
https://youtu.be/4RPh-R3XT8E
256 Hzの音叉によるCochliostema odoratissimumの螺旋状の葯の振動 Vibration of the spiral anthers of Cochliostema odoratissimum induced by a 256 Hz tuning fork
https://youtu.be/fnYgMLHUZPg
256 Hzの音叉によるCochliostema odoratissimumの螺旋状の葯の振動 (拡大) Vibration of the spiral anthers of Cochliostema odoratissimum induced by a 256 Hz tuning fork (magnified)
The staminal cuculli were opened to expose the spiral anthers, and vibrations were applied using a 256 Hz tuning fork. As a result, the anthers were observed to vibrate finely. Because six spiral anthers are densely packed within the staminal cuculli, their resonance may generate the driving force that ejects pollen forcefully.
引用文献 References
De Crane LPR. 2010. Floral Diagrams. (Cambridge Univ. Press: Cambridge).
De Luca PA, Buchmann S, Galen C, Mason AC, and Vallejo-Marín M. 2019. Does body size predict the buzz-pollination frequencies used by bees? Ecol Evol. 9: 4875–4887. https://doi.org/10.1002/ECE3.5092
De Luca PA, Bussière LF, Souto-Vilaros D, Goulson D, Mason AC, and Vallejo-Marín M. 2012. Variability in bumblebee pollination buzzes affects the quantity of pollen released from flowers. Oecologia 172: 805–816. https://doi.org/10.1007/S00442-012-2535-1
Endress PK. 1994. Diversity and Evolutionary Biology of Tropical Flowers (Cambridge Univ. Press: Cambridge).
Evans TM, Faden RB, Simpson MG, Sytsma KJ. 2000. Phylogenetic relationships in the Commelinaceae: I. A cladistic analysis of morphological data. Syst. Bot. 25: 668–691.
Hardy, C.R. and Faden, R.B. 2004. Plowmanianthus, a new genus of Commelinaceae with five new species from tropical america. Syst Bot. 29: 316–333. https://doi.org/10.1600/036364404774195511
Hardy, C. and Stevenson, D. 2000. Development of the gametophytes, flower, and floral vasculature in Cochlioptema odoratissimum (Commelinaceae). Bot J Linn Soc. 134: 131–157.
Mesquita-Neto JN, Blüthgen N, and Schlindwein C. 2018. Flowers with poricidal anthers and their complex interaction networks—Disentangling legitimate pollinators and illegitimate visitors. Funct Ecol. 32: 2321–2332. https://doi.org/10.1111/1365-2435.13204
Vallejo-Marin M and Russell AL. 2024. Harvesting pollen with vibrations: towards an integrative understanding of the proximate and ultimate reasons for buzz pollination. Ann Bot. 133: 379–398. https://doi.org/10.1093/AOB/MCAD189
Tristerix corymbosus: Photo by shmulia, https://www.inaturalist.org/photos/492796211, CC BY-NC Tristerix penduliflorus: Photo by J.R. Kuethe, https://www.inaturalist.org/photos/341912303, CC BY-NC Tristerix longibracteatus: Photo by Joey Santore, https://www.inaturalist.org/photos/613341336, CC BY-NC Tristerix verticillatus: Photo by Thibaud Aronson, https://www.inaturalist.org/photos/354120186, CC BY-SA Tristerix secundus: Photo by sussandro, https://www.inaturalist.org/photos/124997156, CC BY-NC
Tristerix属は13種からなり、アルゼンチン、チリ、ボリビア、ペルー、エクアドル、コロンビアに自生する(POWO)。鮮やかな赤色の花弁を形成し、花序の末端に1個の花を付ける(monad)、子葉が種子から外に出ず、種子内で胚乳から栄養を得るなどの特徴を共有している(Kuijt and Hansen, 2014)。
Tristerix comprises 13 species and is native to Argentina, Chile, Bolivia, Peru, Ecuador, and Colombia (POWO). It shares several characteristic features, including the formation of conspicuous bright red petals, the presence of a single terminal flower in the inflorescence (monad), and cotyledons that do not emerge from the seed but instead obtain nutrients from the endosperm within the seed (Kuijt and Hansen, 2014).
Tristerix corymbosus: Photo by secanosinsequia, https://www.inaturalist.org/photos/597212575, CC BY-NC
Species of Tristerix are parasitic and do not form roots. Instead, they develop a haustorium that grows into the host tissues, from which they absorb water and nutrients (Kuijt 1988). The seeds are covered with a sticky substance, and after being excreted in bird droppings, they adhere to the bark of the host. The seed contains endosperm, and the fused cotyledons do not emerge from the seed but obtain nutrients from the endosperm within the seed. During germination, a haustorium elongates from the seed. The tip of the haustorium forms a haustorial disc, which either closely adheres to the host tissue or penetrates it by breaking down the tissue and then extends internally. The haustorium is hypothesized to be a modified hypocotyl (Bhatnagar and Johri 1983) or root-shoot mosaic (Teixeira-Costa 2021).
Tristerix aphyllus-1: Photo by tomas1309, https://www.inaturalist.org/photos/488472787, CC BY-NC Tristerix aphyllus-2: Photo by Jorge Herreros de Lartundo, https://www.inaturalist.org/photos/346919105, CC BY
Tristerix aphyllusはチリ中部に分布し(Amico et al. 2007)、姉妹種であるT. corymbosus(Amico et al. 2007)を含む他のTristerix属の種と異なる形態を示す。T. aphyllusを除くTristerix属の種は、光合成可能な葉を付けたシュートを形成する半寄生植物であるのに対し、T. aphyllusは栄養シュートを形成せず葉を形成しない全寄生植物である。
T. aphyllusはサボテン科植物に寄生する唯一の寄生被子植物であり(Ossa et al. 2021)、寄主としてLeucostele (ex Trichocereus or Echinopsis)およびEulychniaが知られている(Mauseth 1990, Amico et al. 2007)。
一般に半寄生植物は、炭水化物を光合成によって自ら生産し、吸器を宿主の導管に接続することで水分と無機塩類を得る(Teixeira-Costa 2021)。一方、全寄生のT. aphyllusでは、吸器が導管に加えて篩管にも接続し、宿主篩管細胞から炭水化物、アミノ酸、脂肪酸などの有機物を得ていると考えられている(Mauseth et al. 1985)。
Tristerix aphyllus occurs in central Chile (Amico et al. 2007) and exhibits a morphology distinct from other species of Tristerix, including its sister species Tristerix corymbosus(Amico et al. 2007). Whereas Tristerix species other than T. aphyllus are hemiparasites that produce shoots bearing photosynthetic leaves, T. aphyllus is a holoparasite that forms no vegetative shoots and bears no leaves.
It is the only angiosperm parasitic plant known to parasitize members of the Cactaceae (Ossa et al. 2021). Leucostele (ex Trichocereus or Echinopsis) and Eulychnia are known as hosts (Mauseth 1990; Amico et al. 2007). In general, hemiparasitic plants produce carbohydrates via photosynthesis and obtain water and inorganic nutrients by connecting their haustoria to the host xylem (Teixeira-Costa 2021). In contrast, in the holoparasite T. aphyllus, the haustorium connects not only to the xylem but also to the phloem, and it is thought to obtain organic compounds such as carbohydrates, amino acids, and fatty acids from host phloem cells (Mauseth et al. 1985).
Tristerix aphyllus-3: Photo by Diego Almendras G., https://www.inaturalist.org/photos/335623362, CC BY-NC Tristerix aphyllus-4: Photo by Fernando Useros López, https://www.inaturalist.org/photos/172223499, CC BY-NC Tristerix aphyllus-5: Photo by Lorenzo Vega, https://www.inaturalist.org/photos/113712543, CC BY-NC Tristerix aphyllus-6: Photo by Benito Rosende, https://www.inaturalist.org/photos/28013312, CC BY-NC
吸器は宿主の柔組織に広がり、花期になると、宿主表面から花序を伸長し(左上写真)、分岐し(左下写真)、開花し(右上写真)、結実する(右下写真)。他のTristerix属の種と同様に、ハチドリ媒花である(Amico et al. 2007)。種子は鳥によって散布される(Amico et al. 2007; Ossa et al. 2021)。種子発芽には果皮 (epicarp) が取り除かれることが必要だが、鳥に摂食され消化される必要はない(Mauseth 1985)。
The haustorium spreads within the host’s parenchymatous tissues. During the flowering season, the inflorescence emerges from the host surface (upper left photo), branches (lower left), flowers (upper right), and sets fruit (lower right). As in other species of Tristerix, it is pollinated by hummingbirds (Amico et al. 2007). Seeds are dispersed by birds (Amico et al. 2007; Ossa et al. 2021). For germination, removal of epicarp is required, but ingestion and digestion by birds are not necessary (Mauseth 1985).
Tristerix aphyllus-7: Photo by kavm01, https://www.inaturalist.org/photos/538791181, CC BY-NC Tristerix aphyllus-8: Photo by Francisco Riquelme Tapia, https://www.inaturalist.org/photos/419928062, CC BY-NC
Seeds of Tristerix aphyllus, like those of other species of Tristerix, have cotyledons that do not emerge from the seed but instead obtain nutrients from the endosperm within the seed. At germination, a haustorium bearing a haustorial disc at its tip elongates. However, because T. aphyllus parasitizes cacti with a thick cuticular layer, it invades host tissues using a specialized mode of haustorial penetration.
人工的にLeucostele chiloensis (ex Echinopsis chilensis)に播種した場合、17ヶ月後に花序が出現したとの報告がある(Botto-Mahan 2000)。寄生された宿主は、栄養資源を奪われるため、生育が抑制される(Silva and Martínez de Rio 1996)。
The cuticle of the host columnar cactus (shown as the orange line in the figure) is thick; however, regions bearing stomata form depressions, and the stomata are located beneath these depressions. The surface of the haustorium is densely covered with multiseriate trichomes composed of several cell rows. When the haustorium is positioned over a stoma, these multiseriate trichomes elongate toward the stoma and pass through the opening between the guard cells. Because a substomatal cavity is present, the trichomes can reach the palisade cortex without penetrating the thick-walled, rigid hypodermal layer, and subsequently spread into the cortex palisade, cortex, phloem, and xylem (Mauseth 1985). How the multiseriate trichomes recognize the position of stomata and enter the stomatal cavity remains unknown.
When seeds were artificially sown onto Leucostele chiloensis (ex Echinopsis chilensis), inflorescences were reported to emerge 17 months after infection (Botto-Mahan 2000). Parasitized hosts exhibit reduced growth due to the loss of nutrients (Silva and Martínez de Rio 1996).
引用文献 References
Amico, G.C., Vidal-Russell, R., and Nickrent, D.L. 2007. Phylogenetic relationships and ecological speciation in the mistletoe Tristerix (Loranthaceae): The influence of pollinators, dispersers, and hosts. Amer. J. Bot.94: 558–567.
Bhatnagas, R.P. and Johri, B.M. 1983. Embryology of Loranthaceae. In M. Calder and P. Bernhardt eds., The biology of Mistletoes. Academic Press.
Botto-Mahan C, Medel, R., Ginocchio, R., and Montenegro, G. Factors affecting the circular distribution of the leafless mistletoe Tristerix aphyllus (Loranthaceae) on the cactus Echinopsis chilensis. Revista Chilena de Historia Natural. 2000:73:525–531.
Kuijt, J. 1988. Revision of Tristerix (Loranthaceae). Systematic Botany Monographs, Vol. 19. pp. 1-61.
Kuijt, J. and Hansen, B. 2014. The Families and Genera of Vascular Plants. Vol. XII. Flowering Plants Eudicots. Santalales, Balanophorales. K Kubitzki, ed., Springer.
Mauseth, J.D., Montenegro, G., and Walckowiak, A. M. 1985. Host infection and flower formation by the parasite Tristerixaphyllus (Loranthaceae). Can. J. Bot. 63: 567-581.
Mauseth, J.D. 1990. Morphogenesis in a highly reduced plant: the encophyte of Tristerix aphyllus (Loranthaceae). Bot. Gaz. 151: 348–353.
Ossa, C.G., Aros-Mualin, D., Mujica, M.I., and Pérez, F. 2021. The physiological effect of a holoparasite over a cactus along an environmental gradient. Front. Plant Sci.12: 763446.
POWO. Royal Botanic Gardens, Kew., https://powo.science.kew.org/, accessed: 6 February 2026.
Silva, A. and Martinez del Rio, C. 1996. Effects of the mistletoe Tristerix aphyllus (Loranthaceae) on the reproduction of its cactus host Echinopsis chilensis. Oikos 75: 437-442. Teixeira-Costa, L. 2021. A living bridge between two enemies: haustorium structure and evolution across parasitic flowering plants. Brazilian J. Bot. 44: 165–178.
カテゴリー:5.15.Santalales.ビャクダン目, parasite 寄生植物|サボテンに寄生するマツグミ科ビャクダン目の種 A species of Loranthaceae (order Santalales) parasitic on cacti (family Cactaceae). はコメントを受け付けていません
ハスイモ属はハスイモ elephant’s ear1種からなる(POWO 2025)。ハスイモ(giant elephant ear or Indian taro)の葉は巨大で、1.5 mから3 mにも成長し、葉柄は食用される (Ivancic et al. 2008)。
The genus Leucocasia comprises a single species, Leucocasia gigantea, commonly known as giant elephant ear or Indian taro (POWO 2025). Its leaves are enormous, growing to 1.5 to 3 meters in length, and the petioles are edible (Ivancic et al. 2008).
東南アジア(図の白線の内側)に野生し、日本を含めた地域で栽培されている(POWO 2025)。
It grows wild in Southeast Asia (within the white line on the map) and is cultivated in regions including Japan (POWO 2025).
花序は仏炎苞に覆われ、雌性先熟で、雌花が成熟した1から数日後に雄蕊から花粉が放出される(Ivancic et al. 2008)。雌蕊と雄蕊はそれぞれの成熟期に発熱し、ザセンソウなど、他の温度発生性花序と同じように、花粉媒介昆虫を誘引する(Ivancic et al. 2008)。温度が42度を超える場合もあり、昆虫を追い出す機能もあるのではないかと議論されているが、野外における検証実験が必要である(Ivancic et al. 2008)。
Its inflorescence is enclosed by a spathe, and the plant is protogynous: the pistillate flowers mature first, and pollen is released from the stamens one to several days later (Ivancic et al. 2008). Both the pistils and stamens undergo thermogenesis during their respective maturation phases. As in other thermogenic inflorescences such as Symplocarpus, this heat is thought to attract insect pollinators (Ivancic et al. 2008). Temperatures can exceed 42°C, raising the possibility that the heat may also serve to drive insects away, although this hypothesis requires field testing (Ivancic et al. 2008).
The scientific name Leucocasia gigantea (Blume) Schott is currently used, though the taxonomy has changed over time. Blume (1823), noting its large white spathe, originally classified the species in the genus Caladium (which includes other species with similar spathes) and named it Caladium giganteum Blume.
Later, Schott (1857) emphasized that while Caladium species typically have multi-locular ovaries and anatropous ovules, this species has a nearly unilocular ovary (subunilocular) and orthotropous ovules. He therefore established a new genus, Leucocasia, consisting only of this species, naming it Leucocasia gigantea (Blume) Schott.
その後、Hooker (1893)は、本種の胚珠が基底胎座basal placentationではなく分散した側膜胎座parietal and scattered placentationであると解釈し、多数の胚珠を付けることなど、Colocasia属に類似していることから、Colocasia gigantea (Blume) Hook.f.とした。
Subsequently, Hooker (1893) interpreted the ovules of this species as having parietal and scattered placentation, rather than basal placentation. Due to this and the large number of ovules, which resemble those of Colocasia, he reclassified the species as Colocasia gigantea (Blume) Hook.f.
しかし、核ゲノムのphytochrome C (PHYC)遺伝子、葉緑体ゲノムのtrnL intron、trnL–trnF intergenic spacer、rpl20–rps12 intergenic spacer, trnK/matK regionの解析から、本種は、クワズイモ属Alocasiaの姉妹種となり、サトイモ属Colocasiaとは単系統にならないことがわかった (Nauheimer et al. 2012)。そこで、現在では、Colocasia gigantea (Blume) Hook.f.ではなく、Leucocasia gigantea (Blume)Schottが用いられる。
However, phylogenetic analyses of the nuclear gene phytochrome C (PHYC) and chloroplast genome regions including the trnL intron, trnL–trnF intergenic spacer, rpl20–rps12 intergenic spacer, and trnK/matK region revealed that this species is a sister group to the genus Alocasia, and not monophyletic with Colocasia (Nauheimer et al. 2012). Thus, Leucocasia gigantea (Blume) Schott is now the accepted name rather than Colocasia gigantea (Blume) Hook.f.
インドクワズイモAlocasia macrorrhizos (L.) G.Donもハスイモに似た厚い大きな葉を形成するが、近縁ではなく(Nauheimer et al. 2012)、この形質は平行進化したと考えられる。
Although Alocasia macrorrhizos (L.) G.Don (commonly known as giant taro) also produces large, thick leaves similar to those of Leucocasia, it is not closely related (Nauheimer et al. 2012), and the shared leaf morphology is considered a result of parallel evolution.
1. Ovules are many and parietal —– 2 1. Ovules are few and basal —– Alocasia 2. Plants often exceed 2 m high; Limb of spathe pure white —– Leucocasia 2. Plants are usually less than 2 m high; Limb of spathe is not pure white —– Colocasia
引用文献 References
Blume, C.L., 1823. Catalogus van eenige der merkwaardigste zoo in- als uitheemsche gewassen te vinden iv ,s Lands Plantentuin te Buitenzorg/opgemaakt door C.L. Blume. (Reprinted by the Arnold Arboretum, 1946.)
GBIF 2025. Leucocasia gigantea (Blume) Schott in GBIF Secretariat (2023). GBIF Backbone Taxonomy. Checklist dataset https://doi.org/10.15468/39omei accessed via GBIF.org on 2025-06-14.
Hooker, J.D. 1893. Aroideae. Flora of British India, vol. 6. L. Reeve & Co., London.
Schott, H.W. 1857. Leucocasia, eine Gattung der Colocasinae. Oesterr. Bot. Wochenbl., 7: 33-35.
Ivancic A, Roupsard O, Garcia JQ, Melteras M, Molisale T, Tara S, and Lebot V. 2008. Thermogenesis and flowering biology of Colocasia gigantea, Araceae. J. Plant Res. 121:73–82.
Nauheimer, L., Boyce, P.C., Renner, S.S. 2012. Giant taro and its relatives: A phylogeny of the large genus Alocasia (Araceae) sheds light on Miocene floristic exchange in the Malesian region. Mol. Phylogenet. Evol. 63: 43–51.
POWO (2025). “Plants of the World Online. Facilitated by the Royal Botanic Gardens, Kew. Published on the Internet; https://powo.science.kew.org/. Retrieved 14 June 2025.”
カテゴリー:0.3.0.Leaf 葉, 5.06.02.Alismatales.オモダカ目, Araceae サトイモ科, Ovule 胚珠, Placentation 胎座|巨大な葉を持つハスイモ Leucocasia giganteaがクワズイモ属Alocasiaやサトイモ属Colocasiaと違う点: Differences Between Giant Elephant’s ear Leucocasia gigantea, Which Has Gigantic Leaves, and the Genera Alocasia and Colocasia はコメントを受け付けていません
被子植物は胚発生初期に茎頂分裂組織ができ、その後は、葉腋に新しい茎頂分裂組織ができ、枝となる。葉腋にできる新しい分裂組織(腋芽原基)は、茎頂分裂組織と葉の相互作用によってできる(総説としてWang and Jiao 2018, Nicolas and Laufs 2022)。そのためか、被子植物の99%以上の種で、成長途中に傷害などを受けなければ、葉腋以外に新しい茎頂分裂組織が形成されることはない。マルバマンネングサの場合、葉の無いところから枝(4や7)が出ている点が奇妙である。
The inflorescences of Sedum makinoi have a peculiar structure (Murata 2024 in Japanese). During the vegetative phase, the leaves are arranged in an opposite decussate phyllotaxis (1). When entering the reproductive phase, the apical meristem differentiates into a floral primordium and forms a flower (2). Between the flower and immediately below opposite leaves (3), branches (4) emerge oppositely to the leaves. At the tip of the branch, there is a single leaf (5), and in the axil of this leaf, a flower (6) forms. From between this leaf (5) and the flower (6), branches (7) grow out in the opposite decussate direction relative to the leaf (5). This process is repeated sequentially, forming the curious inflorescence.
In angiosperms, a shoot apical meristem is formed during early embryogenesis, and later, new shoot apical meristems are formed in the leaf axils, giving rise to branches. The new meristem that forms in the leaf axils (axillary meristem primordia) is generated through the interaction between the shoot apical meristem and the leaves (reviewed in Wang and Jiao 2018, Nicolas and Laufs 2022). As a result, in over 99% of angiosperm species, new apical meristems are not formed outside of leaf axils unless the plant is damaged during growth. In the case of Sedum makinoi, it is peculiar that branches (4 and 7) emerge from areas where there are no leaves.
マルバマンネングサの花序形成機構の仮説 Hypothesis on the inflorescence development of Sedum makinoi
The seemingly peculiar morphology of angiosperms can often be explained by anisotropic growth. Anisotropic growth refers to the growth of specific regions of the plant at a rate or manner different from other regions. The unusual inflorescence of Sedum makinoi can be effectively explained by considering that the base region, including the leaf primordium and its axillary meristem, undergoes differential growth as described below.
In regular angiosperms, it appears that the apical region grows almost uniformly (a). However, in Sedum makinoi, the base of the leaf and its axillary meristem primordium elongates more than other regions (b), causing the portion that includes a single leaf primordium (c) and its axillary meristem (d) to extend and forms a branch (e). The leaf primordium develops into a leaf (“C”). The axillary meristem (d) forms opposite leaf primordia, their axillary meristem, and a terminal flower (f) just like in the previous shoot. The base (i) of the leaf primordia (g) and axillary meristem primordia (h) elongates (D). Repeating this process results in the inflorescence morphology characteristic ofSedum makinoi. Although the inflorescence of Sedum makinoi may seem peculiar at first glance, it appears that this morphology could have evolved through a simple modification—anisotropic growth at the base of the leaf primordia and the axillary region. Investigating where anisotropic growth occurs during inflorescence formation would allow us to test this hypothesis.
基礎生物学研究所生物進化研究部門の長谷部光泰が作っている「植物発生進化学」の講義録です。Lecture notes of “Plant Development and Evolution” by Mitsuyasu Hasebe at National Institute for Basic Biology (NIBB), Japan.
間違いがありましたら、mhasebe (at mark) nibb.ac.jpまでご連絡いただければ幸いです。If you find any mistakes, kindly inform me to mhasebe (at mark) nibb.ac.jp