被子植物は胚発生初期に茎頂分裂組織ができ、その後は、葉腋に新しい茎頂分裂組織ができ、枝となる。葉腋にできる新しい分裂組織(腋芽原基)は、茎頂分裂組織と葉の相互作用によってできる(総説としてWang and Jiao 2018, Nicolas and Laufs 2022)。そのためか、被子植物の99%以上の種で、成長途中に傷害などを受けなければ、葉腋以外に新しい茎頂分裂組織が形成されることはない。マルバマンネングサの場合、葉の無いところから枝(4や7)が出ている点が奇妙である。
The inflorescences of Sedum makinoi have a peculiar structure (Murata 2024 in Japanese). During the vegetative phase, the leaves are arranged in an opposite decussate phyllotaxis (1). When entering the reproductive phase, the apical meristem differentiates into a floral primordium and forms a flower (2). Between the flower and immediately below opposite leaves (3), branches (4) emerge oppositely to the leaves. At the tip of the branch, there is a single leaf (5), and in the axil of this leaf, a flower (6) forms. From between this leaf (5) and the flower (6), branches (7) grow out in the opposite decussate direction relative to the leaf (5). This process is repeated sequentially, forming the curious inflorescence.
In angiosperms, a shoot apical meristem is formed during early embryogenesis, and later, new shoot apical meristems are formed in the leaf axils, giving rise to branches. The new meristem that forms in the leaf axils (axillary meristem primordia) is generated through the interaction between the shoot apical meristem and the leaves (reviewed in Wang and Jiao 2018, Nicolas and Laufs 2022). As a result, in over 99% of angiosperm species, new apical meristems are not formed outside of leaf axils unless the plant is damaged during growth. In the case of Sedum makinoi, it is peculiar that branches (4 and 7) emerge from areas where there are no leaves.
マルバマンネングサの花序形成機構の仮説 Hypothesis on the inflorescence development of Sedum makinoi
The seemingly peculiar morphology of angiosperms can often be explained by anisotropic growth. Anisotropic growth refers to the growth of specific regions of the plant at a rate or manner different from other regions. The unusual inflorescence of Sedum makinoi can be effectively explained by considering that the base region, including the leaf primordium and its axillary meristem, undergoes differential growth as described below.
In regular angiosperms, it appears that the apical region grows almost uniformly (a). However, in Sedum makinoi, the base of the leaf and its axillary meristem primordium elongates more than other regions (b), causing the portion that includes a single leaf primordium (c) and its axillary meristem (d) to extend and forms a branch (e). The leaf primordium develops into a leaf (“C”). The axillary meristem (d) forms opposite leaf primordia, their axillary meristem, and a terminal flower (f) just like in the previous shoot. The base (i) of the leaf primordia (g) and axillary meristem primordia (h) elongates (D). Repeating this process results in the inflorescence morphology characteristic ofSedum makinoi. Although the inflorescence of Sedum makinoi may seem peculiar at first glance, it appears that this morphology could have evolved through a simple modification—anisotropic growth at the base of the leaf primordia and the axillary region. Investigating where anisotropic growth occurs during inflorescence formation would allow us to test this hypothesis.
ブドウ科の葉に対生する蔓 Leaf-opposed tendrils in the grape family, Vitaceae
ブドウのシュートを観察すると、葉と対生の位置に蔓(ツル:Tendril)が付いている(Gerrath and Posluszny 2007, 邑田と前田 2022)。このツルのつき方は、他の被子植物では見られず、ブドウ科のツルを作る全ての種に共通であることから(Wen 2007)、ブドウ科の共有派生形質だと考えられる。
When observing grape shoots, tendrils are attached opposite the leaves (Gerrath and Posluszny 2007, 邑田と前田 2022). This attachment pattern of tendrils is not observed in other angiosperms and is common to all tendril-producing species in the grape family (Vitaceae, Wen 2007). Therefore, it is considered a synapomorphy of the Vitaceae.
ブドウ科の葉に対生する蔓は腋芽が変化してできる単軸分枝 Leaf-opposed tendrils of the Vitaceae are formed from axillary meristems as monopodial branching
ブドウ科の4属(ノブドウ属 Ampelopsis、セイシカズラ属 Cissus、ツタ属 Parthenocissus、ブドウ属 Vitis)10種における茎頂表面の落射照明光学顕微鏡や走査電子顕微鏡による観察や、茎頂切片の光学顕微鏡観察から(Bugnon 1964, Millington 1966, Tucker and Hoefert 1968, Shah and Dave 1970, Gerrath and Posluszny 1988, Gerrath and Posluszny 1989a, Gerrath and Posluszny 1989b, Gerrath and Posluszny 1994, Wilson and Posluszny 2003, Gerrath and Posluszny 2007, Timmons et al. 2007) 、もともとの茎頂分裂組織は維持され、腋芽が蔓原基となっていることがわかった(総説としてGerrath and Posluszny 2007)。このような成長様式を単軸分枝と呼び、シロイヌナズナの栄養成長期に見られる分枝様式と同じである。シロイヌナズナの茎は伸長しないが、タバコなどの被子植物の茎は、腋芽と次の葉の間の部分が伸長する。ブドウ科で変わっている点は、腋芽とその蓋葉(腋芽に伴っている葉)原基(図のL2)の間の組織(黄色い部分)が伸長することで、次の葉原基(L1)と対生の位置に蔓が配置する点である。したがって、葉に対生する蔓というブドウ科に特有の体制は、茎が偏差成長することによって進化したと考えられる。
From epi-illumination light microscopy and scanning electron microscopy of the shoot apical meristem and light microscopy of the sections in ten species across four genera of the Vitaceae (Ampelopsis, Cissus, Parthenocissus, and Vitis) (Bugnon 1964, Millington 1966, Tucker and Hoefert 1968, Shah and Dave 1970, Gerrath and Posluszny 1988, Gerrath and Posluszny 1989a, Gerrath and Posluszny 1989b, Gerrath and Posluszny 1994, Wilson and Posluszny 2003, Gerrath and Posluszny 2007, Timmons et al. 2007), it has been found that the original shoot apical meristem is maintained, and the axillary meristem forms the tendril primordium (reviewed in Gerrath and Posluszny 2007). This type of growth is called monopodial branching, which is the same branching pattern observed during the vegetative growth phase of Arabidopsis thaliana. While the stem of Arabidopsis does not elongate, in other angiosperms like tobacco, the region between the axillary meristem and the next leaf primordium elongates. The unique characteristic of the Vitaceae is that the tissue (yellow part in the figure) between the axillary meristem (Tendril primordium) and the primordium of subtending leaf (L2 in the figure) elongates, positioning the tendril opposite the next leaf primordium (L1). Therefore, it is considered that the tendril arrangement opposite the leaves, which is a unique trait of the Vitaceae, evolved through anisotrophic stem growth.
Rhoicissus digitataの蔓は仮軸分枝でできる可能性がある Tendrils of Rhoicissus digitata appear to be formed sympodially
一方、Rhoicissus digitataでは、蔓原基ができるころの茎頂で、蔓原基の方が茎頂原基よりも大きいことから、もともとの茎頂分裂組織が蔓原基へと変化し、図のL1の腋芽が新しい茎頂分裂組織を形成している(仮軸分枝と呼ぶ)のではないかと報告された(Gerrath et al. 1998, Gerrath and Posluszny 2007)。また、同属のR. rhomboideaでは、茎頂分裂組織と蔓原基の大きさがほぼ同じである(Gerrath et al. 1998, Gerrath and Posluszny 2007)。シロイヌナズナやイネの腋芽発生初期と茎頂では、発現する遺伝子が異なっており(総説として、Wang and Jiao 2018, Nicolas and Laufs 2022)、遺伝子の発現様式を発生段階を追って調べれば、どこが茎頂でどこが腋芽か、茎頂から腋芽への変化がおきているかなどを調べることが可能である。従来の外部形態や切片による細胞形態による観察に、遺伝子発現様式を加えた研究を行うことで、ブドウ科の蔓がどのように形成されるかがよりはっきりと推定できるだろう。
On the other hand, in Rhoicissus digitata, it has been reported that, at the shoot apex when the tendril primordium forms, the tendril primordium is larger than the shoot apical meristem, suggesting that the original shoot apical meristem transforms into the tendril primordium, and the axillary meristem of L1 forms a new shoot apical meristem (referred to as sympodial branching) (Gerrath et al. 1998, Gerrath and Posluszny 2007). In the related species R. rhomboidea, the shoot apical meristem and the tendril primordium are almost the same size (Gerrath et al. 1998, Gerrath and Posluszny 2007). In Arabidopsis thaliana and rice, different sets of genes are expressed at the shoot apical meristem and the initial stage of axillary meristem formation (reviewd in Wang and Jiao 2018, Nicolas and Laufs 2022), and by investigating the gene expression patterns over developmental stages, it is possible to determine the boundary between the shoot apex and the axillary bud and to investigate whether a transformation from shoot apical meristem to axillary meristem is occurring. By combining studies of traditional external morphology and cell morphology in sections with gene expression patterns, it would become clearer how tendrils in the Vitaceae are formed.
References:
Bugnon, F. 1964. Sur les modes de ramification de quelques inflorescences dont la significantion est controversée. Bull. Sci. Bot. Fr. Mémoir. 111: 101-124.
Gerrath, J.M. and Posluszny, U. 1988. Morphological and anatomical development in the Vitaceae. I. Vegetative development in Vitis riparia. Can. J. Bot. 66: 209-224.
Gerrath, J.M. and Posluszny, U. 1989a. Morphological and anatomical development in the Vitaceae. III. Vegetative development in Parthenocissus inserta. Can. J. Bot. 67: 803-816.
Gerrath, J.M. and Posluszny, U. 1989b. Morphological and anatomical development in the Vitaceae. V. Vegetative and floral development in Ampelopsis brevipedunculata. Can. J. Bot. 67: 2371-2386.
Gerrath, J.M. and Posluszny, U. 1994. Morphological and anatomical development in the Vitaceae. VI. Cissus antarctica. Can. J. Bot. 72: 635-643.
Gerrath, J.M. and Posluszny, U. 2007. Shoot architecture in the Vitaceae. 2007. Can. J. Bot. 85: 691-700.
Gerrath, J.M., Lacroix, C.R., and Posluszny, U. 1998. Phyllotaxis in the Vitaceae. In Symmetry in Plants. Ed. R.V. Jean and D. Barabé. World Scientific Publication, Singapore. pp. 89-107.
Millington, W.F. 1963. Long shoots, short shoots and tendril shoots in the morphogenesis of Parthenocissus. Amer. J. Bot. 53: 74-81.
邑田仁、前田綾子. 2022. ブドウ科植物のシュートの構造. J. Japanese Bot. 97: 33-50.
Nicolas, A. and Laufs, P. 2022. Meristem initiation and de novo stem cell formation. Front. Plant Sci. 13: 891228.
Shah, J.J. and Dave, Y.S. 1970. Morpho-histogenetic studies on Tendrils of Vitaceae. Amer. J. Bot. 57: 363-373.
Timmons, S.A., Posluszny, U., and Gerrath, J.M. 2007. Morphological and anatomical development in the Vitaceae. X. Comparative ontogeny and phylogenetic implications of Cissus quadrangularis L. Can. J. Bot. 85: 860-872.
Tucker, S.C. and Hoefert, L.L. 1968. Ontogeny of the tendril in Vitis viniferra. Amer. J. Bot. 55: 1110-196.
Wang, Y. and Jiao, Y. 2018. Axillary meristem initiation — a way to branch out. Curr. Opin. Plant Biol. 41: 61–66.
Wen, J. Vitaceae. 467-479. In K. Kubitzki (ed.) 2007. The Families and Genera of Vascular Plants. IX. Springer. Wilson, T. and Posluszny, U. 2003. Complex tendril branching in two species of Parthenocissus: implications for the vitaceous shoot architecture. Can. J. Bot. 81: 587-597.
カテゴリー:tendril 蔓, 未分類|ブドウ科の葉に対生する蔓 Leaf-opposed tendrils in the grape family Vitaceae はコメントを受け付けていません
The Venus flytrap genus, Dionaea, captures small animals using modified leaves that act as a trap, obtaining nutrients to thrive in nutrient-poor environments. It is a sister genus to the sundew genus, which is also a carnivorous plant, and consists of a single species, Dionaea muscipula, making it a monotypic genus.
米国ノースカロライナ州のハエトリソウの自生地 Locality of the Venus flytrap Dionaea muscipula near Wilmington, North Carolina, USA
ハエトリソウは米国のノースカロライナ州とサウスカロライナ州の海岸線の沿った約340 kmの範囲に分布していたが、土地開発や採集によって現在では限られた国立公園や私有地などで見られるだけである(Bailey and McPherson 2012, Juniper et al. 1989, Roberts and Oosting 1958)。地図は次のサイトより引用:https://www.freemap.jp/free.html
The Venus flytrap was distributed along a roughly 340 km stretch of the coastal region of North Carolina and South Carolina in the United States, but due to land development and poaching, it can now only be found in limited areas such as national parks and private lands (Bailey and McPherson 2012, Juniper et al. 1989, Roberts and Oosting 1958). The map was sited from https://www.freemap.jp/free.html
米国ノースカロライナ州のハエトリソウの自生地 Locality of the Venus flytrap Dionaea muscipula near Wilmington, North Carolina, USA
ハエトリソウは、山火事が数年おきにおこる場所に生育する。地下の鱗茎は表面を焼く程度の山火事には耐性があり、上部を覆っていた草が焼き払われると元気に生育する。山火事の後には約10%開花個体が増えるが、数年して周りをイネ科、カヤツリグサ科、イグサ科の草本が覆うようになると開花数が減る(Bailey and McPherson 2012, Roberts and oosting 1958)。
The Venus flytrap grows in areas where wildfires occur every few years. The underground bulb is resistant to wildfires that burn only the surface, and it thrives once the grasses covering the top are burned away. After a wildfire, the number of flowering individuals increases by about 10%, but as grasses from the Poaceae, Cyperaceae, and Juncaceae families cover the area over the years, the number of flowering plants decreases (Bailey and McPherson 2012, Roberts and Oosting 1958).
ハエトリソウの形態 Morphology of the Venus flytrap Dionaea muscipula
The leaves of the Venus flytrap have their leaf blades modified into traps, while the petioles grow broadly and take on the appearance of ordinary leaf blades (A). The stem runs horizontally near the surface of soil, and because the internodes between the leaves are short, the leaves form a rosette. The base of the petioles thickens with soft tissue, and the underground part becomes bulbous. (C) is an image of a cross-section of (B), cut along the white dotted line, taken from the shoot apex side of the stem. You can see that the petioles and roots emerge from the thin stem (enclosed in the black dotted line).
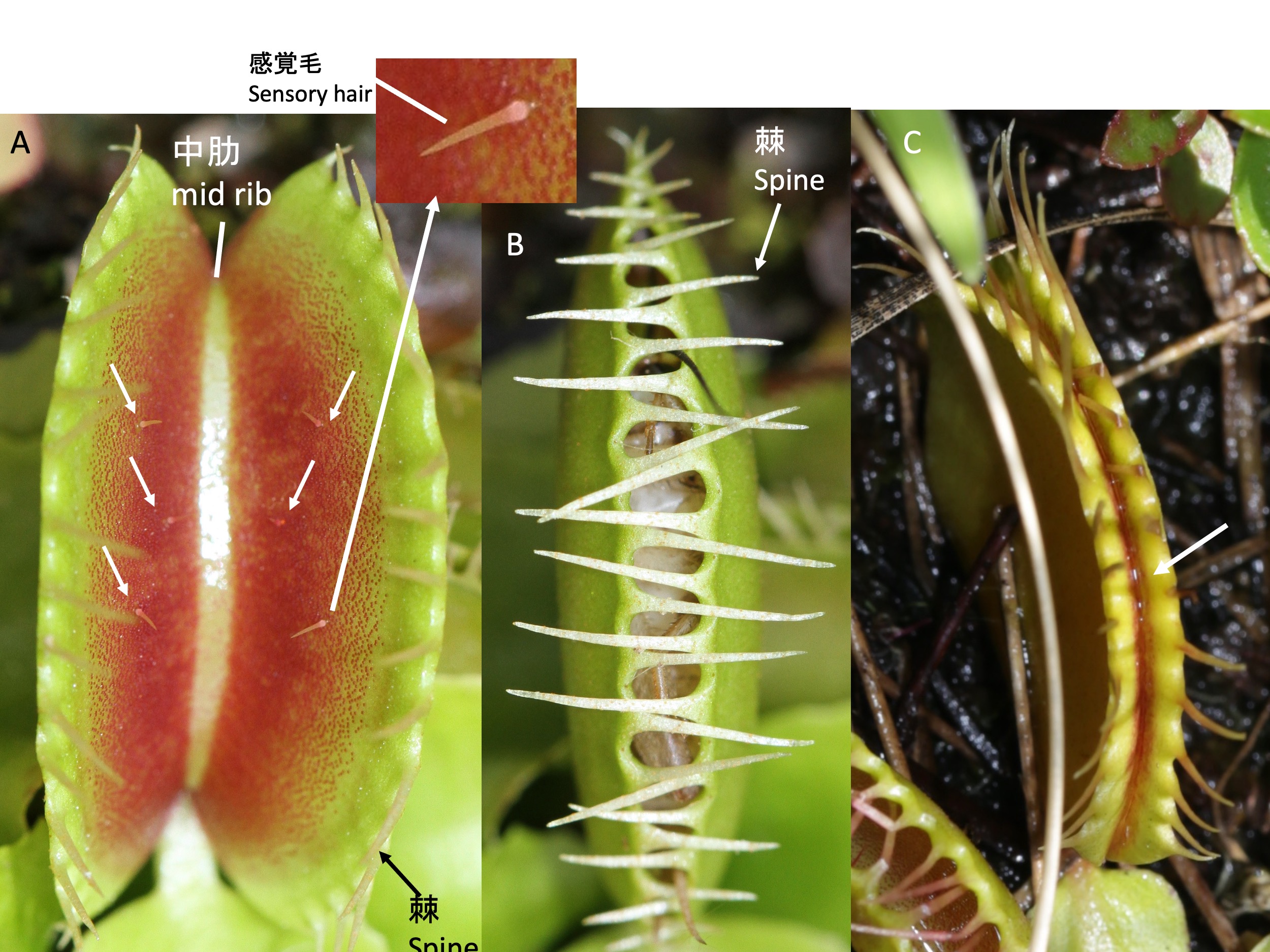
ハエトリソウの捕虫器の形態 Trap morphology of the Venus flytrap Dionaea muscipula
Spines form along the edges (the leaf margin) of the trap (A), and typically six sensory hairs (white arrows in [A]) develop on its surface. When the sensory hairs receive two successive touch stimuli, the trap closes, capturing the prey ([B] and Movie). After the trap closes, if the prey struggles and further stimulates the sensory hairs, the leaf blade bends backward, pressing the prey, and the area just inside the leaf margin seals tightly. The stimulation of the sensory hairs induces the secretion of digestive fluids, and the inner side of the tightly sealed leaf becomes filled with the digestive fluid (the white arrow in [C] indicates where the leaked digestive fluid is visible).
ハエトリソウの捕虫器は感覚毛を2回刺激すると閉じる(須田啓撮影) The trap of the Venus flytrap Dionaea muscipula close with two touch stimuli by Dr. Hiraku Suda
Macfarlane(1902)は、ハエトリソウの感覚毛を1回だけ刺激しても葉が閉じないけれど、20秒間隔で2回刺激すると閉じることを発見した。このことは、ハエトリソウが1回目の刺激を記憶していることを意味している。さらに、1回目と2回目の刺激の間隔を30秒に延ばすと葉が閉じないことから、記憶は30秒ほどしかもたないことにも気づいた。そして、BrownとSharpは記憶時間が温度に依存することに気づいた(Brown and Sharp, 1910)。このことは、記憶が化学反応によっていることを示唆している。Brown (1916)はより定量的な実験を行い、10枚の葉に33秒間隔で2回の刺激を与えると、ほぼ全ての葉が閉じるが、67秒間隔だと平均2.75回の刺激が必要であると報告し、この実験に基づき、ハエトリソウは約30秒の記憶があると記載されることが多い。また、葉が2回の刺激で閉じる仕組みは、落ち葉、小石、雨などの獲物以外の物が偶然あたっても葉を閉じない点で適応的だと考えられている(Poppinga et al., 2018)。特に、雨は数日おきに降るので、雨粒にあたって葉が閉じないようにすることは、エネルギーの損失を防ぐ上で重要だと考えられる。栽培しているハエトリソウにジョウロで水をかけても葉が閉じない。よく見ると、1粒目の雨粒があたると、表面張力によって、水が感覚毛の周りを覆ってしまう。なので、2粒目以降の雨粒があたっても刺激が弱くなっているようである。
植物は、外界からのさまざまな刺激を記憶することが知られている(Taiz et al. 2022)。例えば、乾燥や高温などへの順応、病気にかかると病害抵抗性が強くなる植物免疫、冬を記憶して春に花が咲く春化などである。これらの数日以上の長期間の記憶に対し、ハエトリソウが持つ短期記憶は、植物には稀であることから興味を持たれ、多くの研究が行われてきた。
Here I introduce research on memory using the Venus flytrap (partially modified and cited from Hasebe 2023 [in Japanese]). You might wonder, “What does memory mean in plants?”. Our memory is governed by the brain. So, does this mean that plants, which lack a brain, cannot have memory? In 1902, Macfarlane discovered that while stimulating the sensory hairs of the Venus flytrap once does not cause the trap to close, two stimuli given 20 seconds apart will. This suggests that the Venus flytrap “remembers” the first stimulus. Furthermore, when the interval between the first and second stimulus is extended to 30 seconds, the trap does not close, indicating that this memory only lasts for about 30 seconds. Brown and Sharp later found that the duration of memory depends on temperature (Brown and Sharp, 1910), which implies that the memory is governed by chemical reactions. In 1916, Brown conducted more quantitative experiments, reporting that nearly all of 10 traps closed when stimulated twice at 33-second intervals, but at 67-second intervals, an average of 2.75 stimuli were needed. Based on this experiment, it is often stated that the Venus flytrap has a memory of around 30 seconds. Additionally, the mechanism by which the trap closes after two stimuli is considered adaptive because it prevents the trap from closing accidentally due to non-prey objects like leaves, pebbles, or rain (Poppinga et al., 2018). Rain, in particular, falls only every few days, so preventing the trap from closing in response to raindrops is important for conserving energy. If you water a cultivated Venus flytrap with a watering can, the trap does not close. On closer inspection, when the first raindrop hits, surface tension causes water to cover the area around the sensory hairs. Therefore, subsequent raindrops appear to provide a weaker stimulus.
It is known that plants can “memorize” various environmental stimuli (Taiz et al., 2022). Examples include acclimation to drought or high temperatures, plant immunity that strengthens after exposure to disease, and vernalization, where plants memorize winter and bloom in spring. While these examples are forms of long-term memory, lasting days or more, the short-term memory of the Venus flytrap is rare in plants and has thus attracted much research attention.
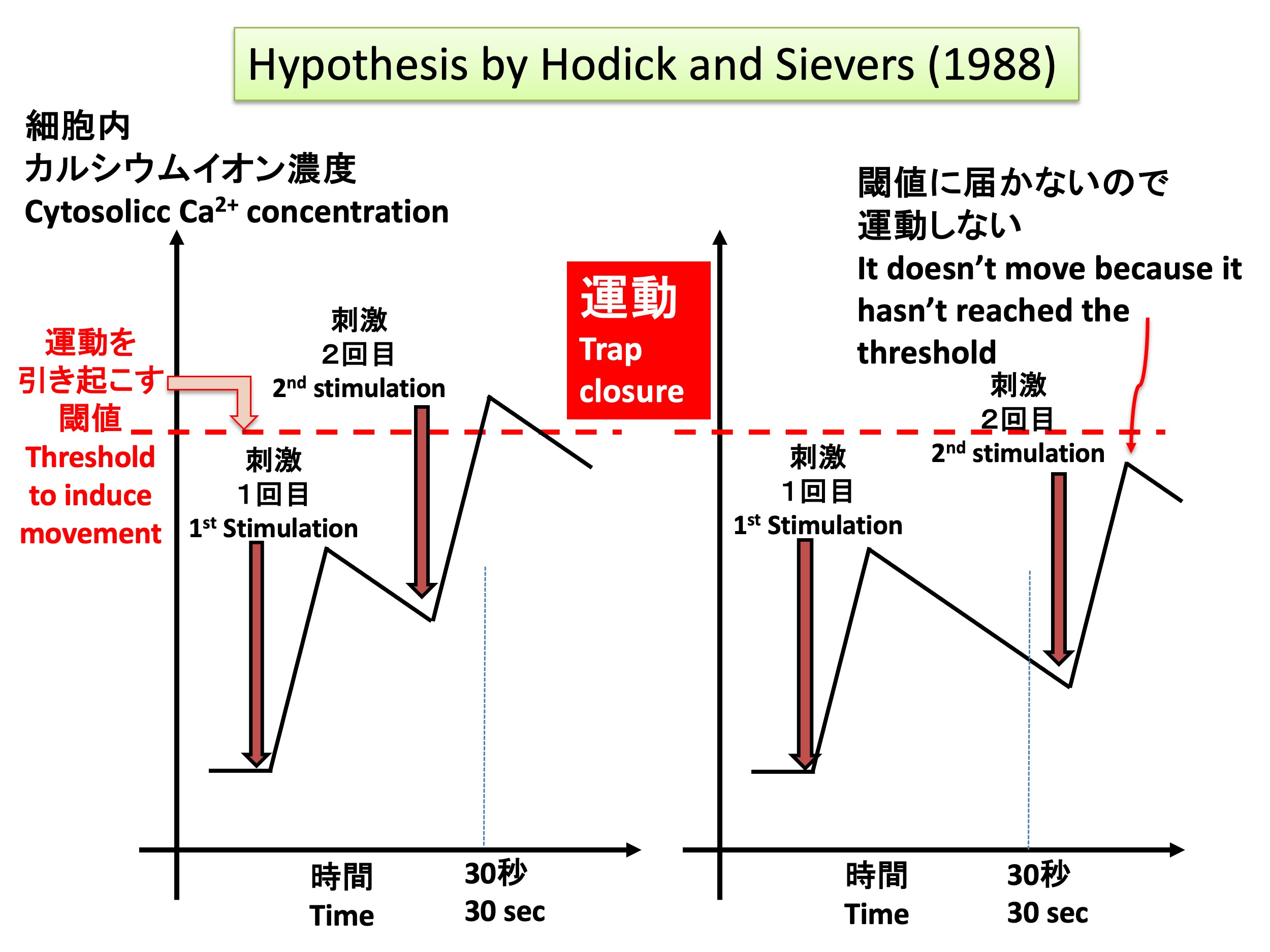
ハエトリソウの記憶の仕組みについてのHodickとSievers (1988)の仮説 Hypothesis of trap movement of the Venus flytrap Dionaea muscipula by Hodick and Sievers (1988)
感覚毛を刺激すると、神経細胞のように、活動電位と呼ばれる一過性の電位変化がおこる(Burdon-Sanderson, 1873)。活動電位は葉が閉じる前に伝搬し(DiPalma et al., 1961)、感覚毛に刺激を与えると、感覚毛から葉全体に同心円状に活動電位が伝搬する(Sibaoka, 1966)。これらのことから、感覚毛から葉に広がる活動電位が刺激となり、なんらかの仕組みを駆動し、運動が引き起こされると考えられるようになった。
動物細胞では、活動電位が細胞膜にあるカルシウムチャネル(特定の条件によってカルシウムイオンを通すタンパク質)を制御することで、細胞質のカルシウムイオン濃度が変化する。カルシウムイオンは二次情報伝達物質として機能し、リン酸化などを介して他のタンパク質の機能を制御することで、細胞に変化を引き起こす。植物でも、巨大細胞を持つホシツリモを用いた研究から、活動電位によってカルシウムチャネルが活性化され、細胞外から細胞内へカルシウムイオンが流入する(Shiina and Tazawa, 1987)。このことから、植物細胞においても、動物細胞同様に、カルシウムが二次情報伝達物質として機能すると考えられるようになり、実際、ミルフラスコモとミナミシャジクモにおいて、活動電位によって細胞質のカルシウムイオン濃度が上昇し、原形質流動が停止することが示された(Hayama et al., 1979)。
When the sensory hairs are stimulated, a transient change in electrical potential called an action potential occurs, similar to what happens in nerve cells (Burdon-Sanderson, 1873). The action potential propagates before the trap closes (DiPalma et al., 1961). When the sensory hairs are stimulated, the action potential spreads concentrically from the hairs throughout the entire leaf (Sibaoka, 1966). From these findings, it became understood that the action potential spreading from the sensory hairs to the leaf serves as a stimulus that triggers some mechanism, causing the movement.
In animal cells, action potentials regulate calcium channels (proteins that allow calcium ions to pass under certain conditions) located in the cell (plasma) membrane, changing the concentration of calcium ions in the cytoplasm. Calcium ions function as secondary messengers, regulating the function of other proteins through processes like phosphorylation, thereby inducing changes in the cell. Similarly, in plants, studies using Nitellaflexilis, a giant-celled green algae, have shown that action potentials activate calcium channels, allowing calcium ions to flow from outside the cell into the cell (Shiina and Tazawa, 1987). This led to the understanding that calcium functions as a secondary messenger in plant cells, just as it does in animal cells. Indeed, in Chara corallina and Nitellopsisobtusa, it was shown that action potentials increase cytoplasmic calcium ion concentrations, which in turn stop cytoplasmic streaming (Hayama et al., 1979).
Hodick and Sievers (1988) proposed a hypothesis that changes in calcium ion concentrations might explain the memory phenomenon, based on these studies and their own research into the mechanism behind action potentials in the Venus flytrap (see figure above). Their hypothesis includes the following points: 1) Stimulation of the sensory hairs generates an action potential, which raises the intracellular calcium ion concentration. 2) When calcium ions or molecules regulated by calcium accumulate beyond a threshold, the trap closes. 3) A single stimulus does not raise the calcium ion concentration above the threshold; at least two stimuli are required. 4) After the calcium ion concentration rises with the first stimulus, it gradually decreases, and after more than 30 seconds, a second stimulus cannot raise the calcium ion concentration above the threshold.
However, in order to test Hodick and Sievers’ hypothesis, it is necessary to observe how the intracellular calcium ion concentration changes. Unfortunately, no method for detecting calcium ion concentrations inside Venus flytrap cells existed, and for over 30 years after the hypothesis was proposed, it remained untested.
ハエトリソウの葉のカルシウムイオン濃度変化を可視化した写真。カルシウム濃度が上がると赤色が濃くなり、さらに濃くなると黄色になる (Suda et al. 2020)。A photo visualizing the changes in calcium ion concentration in the Venus flytrap Dionaea muscipula leaf. As the calcium concentration increases, the red color becomes deeper, and when it increases further, it turns yellow (Suda et al. 2020).Di
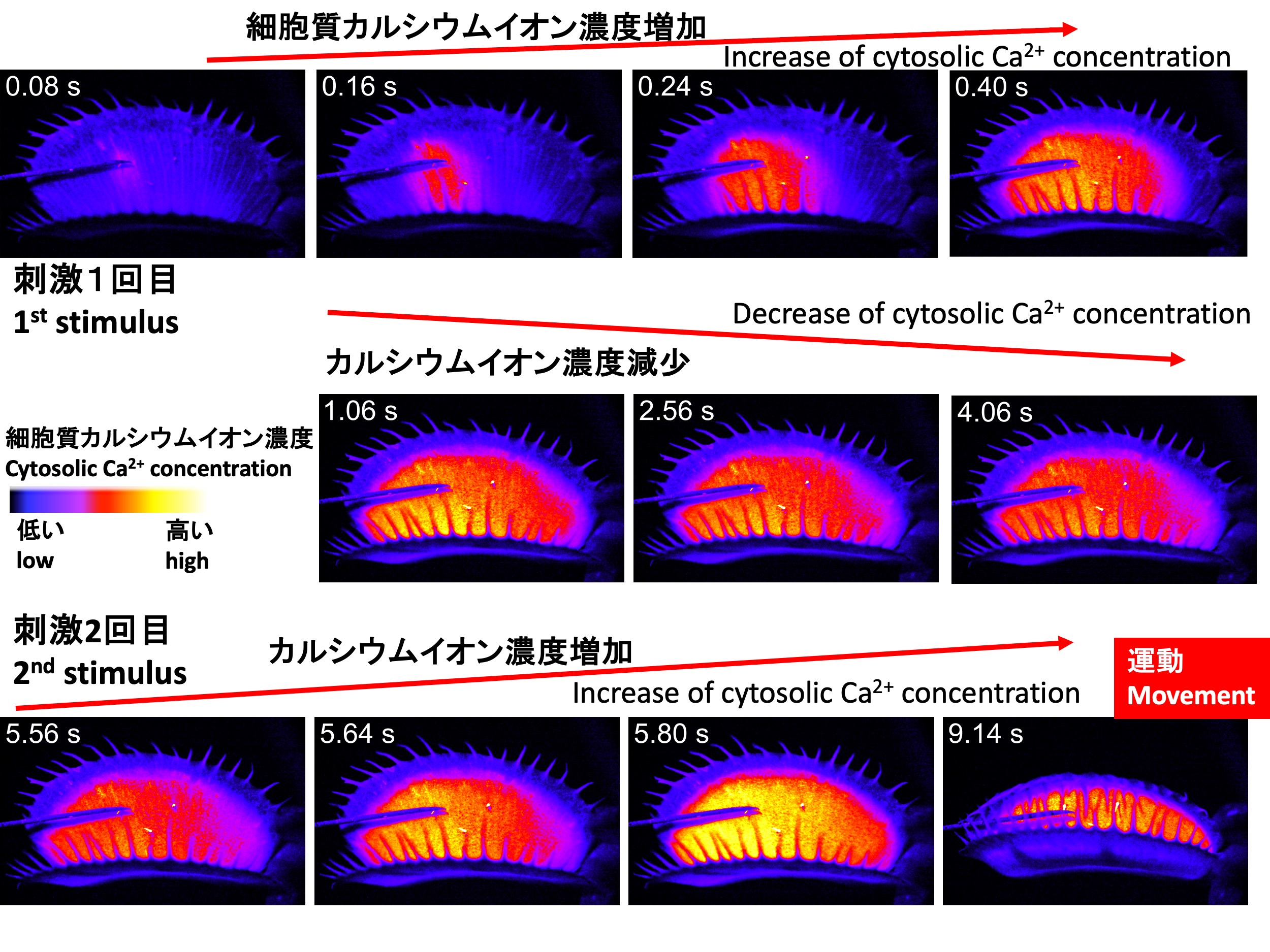
細胞内のカルシウムイオン濃度を検出するには、カルシウムイオンと結合すると光を発するようなセンサータンパク質をハエトリソウで働かせることが必要である。そのためには、センサータンパク質の遺伝子をハエトリソウに導入することが必要である。 そこで我々の研究グループの須田啓は、ハエトリソウにカルシウムイオンと結合すると緑色蛍光を出すGCaMP6fタンパク質をコードする遺伝子を導入し、遺伝子組換えハエトリソウを作出した。そして、感覚毛を1回刺激すると、感覚毛から葉全体に同心円状に緑色の蛍光が広がることを発見した(上図上段)。そして、広がった後、蛍光は時間がたつにつれ、徐々に減少した(上図中段)。2回目の刺激を与えると、1回目よりも強い蛍光が広がり、葉は閉じた(上図下段)(Suda et al., 2020)。
To detect intracellular calcium ion concentrations, it is necessary to use a sensor protein that emits light when it binds to calcium ions. This requires introducing the gene encoding the sensor protein into the Venus flytrap. Therefore, Dr. Hiraku Suda from our research group introduced a gene encoding the GCaMP6f protein, which emits green fluorescence when it binds to calcium ions, into the Venus flytrap, creating a genetically modified Venus flytrap, after mostly three years‘ efforts.
Upon stimulating the sensory hairs once, Suda discovered that fluorescence spreads concentrically from the sensory hairs across the entire leaf (imagens in the top row). After spreading, the fluorescence gradually decreased over time (images in the middle row). When a second stimulus was applied, a stronger fluorescence than after the first stimulus was observed, and the leaf closed (images in the bottom row) (Suda et al., 2020).
ハエトリソウの感覚毛を接触刺激した時の細胞質カルシウムイオン濃度変化(須田啓撮影:Suda et al. 2020) Cytosolic calcium ion dynamics of the Venus flytrap Dionaea muscipula with mechanical stimuli to the sensory hair (Taken by Dr. Hiraku Suda: Suda et al. 2020)
In this video, it is shown that if more than 30 seconds pass after the first stimulus, the trap does not close upon the second stimulus, but if the second stimulus is given within 30 seconds, the trap closes. You can observe that the fluorescence, corresponding to the intracellular calcium ion concentration, gradually weakens over time after the first stimulus.
As a result of the research that began with Burdon-Sanderson’s measurement of action potentials in 1873, it is highly likely that the hypothesis of Hodick and Sievers (1988) is correct, and that the Venus flytrap’s memory can be explained by changes in calcium ion concentration. However, this experiment only shows that calcium ion concentration changes are “correlated” with movement and memory, not that calcium ions are actually functioning as the causal factor. For instance, it is possible that the action potential controls some unknown substance entirely different from calcium ions, and that this substance shows dynamics similar to calcium ions.
To demonstrate a causal relationship, it would be necessary to observe how movement and memory change in vivo by manipulating only the calcium ion concentration without altering the action potential or other substances. However, since calcium ions are involved in generating action potentials and both action potentials and calcium ions regulate various proteins, this experiment would be difficult. Therefore, future research needs to clarify how action potentials and intracellular calcium ion changes are regulated by proteins or other mechanisms, and how intracellular calcium ions induce movement by chemically modifying certain proteins.
Bailey T, McPherson S. 2012. Dionaea. Poole, Dorset, England: Redfern Natural History Productions Ltd.
Brown WH. 1916. The mechanism of movement and the duration of the effect of stimulation in the leaves of Dionaea. Am. J. Bot.3: 68–90.
Brown WH, Sharp LW. 1910. The closing response in Dionaea. Bot. Gaz.49: 290–302.
Burdon-Sanderson JS. 1873. Note on the electrical phenomena which accompany stimulation of the leaf of Dionaea muscipula. Phil. Trans. Roy Soc. London164: 495.
DiPalma JR, Mohl R, Best Jr W. 1961. Action potential and contraction of Dionaea muscipula. Science133: 878–879.
Hayama T, Shimmen T, Tazawa M. 1979. Participation of Ca2+ in cessation of cytoplasmic streaming induced by membrane excitation in Characeae internodal cells. Protoplasma99: 305–321.
Hodick D, Sievers A. 1988. The action potential of Dionaea muscipula Ellis. Planta174: 8–18.
Juniper BE, Robins RJ, Joel DM. 1989. The Carnivorous Plants. London, Uk: Academic Press.
Macfarlane JM. 1902. Contribution to the history of Dionaea muscipula Ellis. Contr. Bot. Lab. Univ. Pennsylvania1: 7–44.
Roberts PR, Oosting HJ. 1958. Responses of Venus Fly Trap (Dionaea muscipula) to Factors Involved in Its Endemism. Ecological Monographs28: 193–218.
Shiina T, Tazawa M. 1987. Demonstration and characterization of Ca2+ channel in tonoplast-free cells of Nitellopsis obtusa. J. Membr. Biol.96: 263–276.
Sibaoka T. 1966. Action potentials in plant organs. Symp. Soc. Exp. Biol.20: 49–73.
Suda H, Mano H, Toyota M, Fukushima K, Mimura T, Tsutsui I, Hedrich R, Tamada Y, Hasebe M. 2020. Calcium dynamics during trap closure visualized in transgenic Venus flytrap. Nature Plants6: 1219–1224.
Taiz L, Miller IM, Murphy A, Zieger E. 2022. Plant Physiology and Development, Seventh Edition. Sinauer Associates Inc.
長谷部光泰(2023) 「食虫植物 多様性と進化」 裳華房
カテゴリー:未分類|ハエトリソウの記憶 Memory of the Venus flytrap はコメントを受け付けていません
育種家のNさんから、Sarracenia purpurea form. heterophylla(以降サラセニアと略す)に1枚だけ奇妙な葉が付いているのに気づいたとのメイルをいただいた。お送りいただいた写真では、同じ株の他の葉は正常なので、突然変異体ではなく、この葉の発生途中になんらかの異常が生じたのではないかと考えられた。奇形葉は、普段は発生しないけれども、なんらかの刺激で形成されうるということは、将来、遺伝的変化でそのような形態が進化する可能性もあり、捕虫葉の進化可能性evolvabilityを考える上で興味深い(Fukushima et al. 2021)。ぜひ観察がしてみたく、お願いしたところ、早々に宅急便が到着した。Nさんに心より御礼申し上げます。捕虫葉を上から覗くと、捕虫葉内面向軸側(茎に近い側)から突起が伸びだしている(Aの白色矢印)。捕虫葉を縦切りにして背側から見ると(B)、突起は口の付近から基部まで繋がっている。横断面を見ると(C)、曲がったキールのように見える。
Ms. N-san, a plant breeder, sent me an e-mail saying that she had noticed one strange leaf on Sarracenia purpurea form. heterophylla (hereafter called Sarracenia). Since the other leaves on the same plant were normal in the photo she sent, I thought that it was not a mutant, but that some kind of abnormality must have occurred during the development of this leaf. The speculation that malformed leaves do not normally occur but can be formed by some stimuli is interesting in considering the evolvability of pitcher leaves (Fukushima et al. 2021), since such a morphology may evolve in the future due to genetic changes. I would very much like to observe it, so I asked her to send me, and the courier arrived in a few days. Thank you very much for N-san! Looking at the pitcher leaf from above, a protuberance is seen extending out from the adaxial inner side of the pitcher leaf (the side close to the stem) (white arrow in A). When the leaf is longitudinally cut and viewed from the abaxial (dorsal) side (B), the protuberance is connected from the mouth to the base. In transverse section (C), the protuberance looks like a curved keel.
The inner surface (A) is not flat, but has a structure with three separate tips (C-E). In C, area of the white dotted line in (B) is enlarged. The structure has three rows of vertical edges (purple, blue, and red stars in [C]). The corresponding area of each star is written on the transverse section in (D), and a schematic diagram is shown in (E). The structure of the keel is such that the tip of the keel is split into three forked pieces.
突起がどのようにできたかを考えるために、まず、通常の捕虫葉の発生過程を見てみよう。(A)から(E)はSarracenia purpurea ssp. venosa var. burkiiの捕虫葉の発生過程を走査電子顕微鏡で撮影したものである(Fukushima et al. 2015)。最初は平面葉のように平面上に発生するが(A、B)、100ミクロンほどになると向軸側に窪みができ、成長するにつれ、窪みが深くなる。(F)は(E)よりも大きくなった葉原基の縦断面である。
To consider how the protuberances were formed, let us first look at the developmental process of a normal pitcher leaf. (A) to (E) are scanning electron microscopic images of the developmental process of a pitcher leaf of Sarracenia purpurea ssp. venosa var. burkii (Fukushima et al. 2015). Initially, they develop on a flat surface like flat leaves (A, B), but at about 100 µm, it is adaxially concaved, and as they grow, further concaved deeper. (F) is a longitudinal section of a leaf primordium that is larger than (E).
窪みは、細胞の分裂方向が変化することによってできることがわかってきた。通常の植物の平面葉では、葉の表面に対して垂直方向の細胞分裂(垂層分裂:Aの断面図の赤線)が多く起こることで、葉は平面状に広がって成長する(Esau 1977)。サラセニアの葉では、先端側は平面葉と同じように垂層分裂(Bの断面図の赤線)がおきるが、基部側では、葉の表面と平行は細胞分裂(並層分裂:Bの断面図の水色線)を起こし、表面方向に出っ張って成長する。例えるならば(C)、サラセニアの葉の発生は、Tシャツを着てみぞおちのあたりを上側に引っ張ると、みぞおちのあたりに窪み(Cの白色矢印)ができるのと同じ仕組みである。そして、並層分裂が引き続くことによって、キールが形成される。どうして基部向軸側のみ並層分裂が起こるのかはわかっていない。オオバナイトタヌキモの捕虫葉形成についても研究が進んでいるが(Whitewood et al. 2020)、キールが形成されないので、サラセニアとは異なった仕組みだと考えられる。図は長谷部(2020)より引用。
It is known that concave is formed by changes in the direction of cell division (Fukushima et a. 2015). In the planar leaves of normal plants, cells in the subepidermal layers divide perpendicularly to the leaf surface (anticlinal cell divisions: red lines in the cross section in [A]), causing the leaf to expand and grow in a planar shape (Esau 1977). In Sarracenia leaves, the distal part of leaf undergoes anticlinal cell division (red line in cross section in [B]) as in planar leaves, but at the proximal part, cells divide parallel to the leaf surface (periclinal cell divisions: light blue line in cross section in [B]). The development of Sarracenia leaf is similar to the formation of a depression (white arrow in C) around the midsection of a T-shirt when the shirt is pulled upward around the midsection. The keel is then formed by continued periclinal cell divisions in the adaxial subepidermal cell layers. It is not known why the periclinal cell divisions are induced in the adaxial side. The formation of the bladder carnivorous leaf in Utricularia gibba has also been well studied (Whitewood et al. 2020), but since the keel is not formed, the mechanism is thought to be different from that of Sarracenia. Figure is cited from 長谷部 (2020).
通常の捕虫葉では、キールは向軸側の表皮に近い細胞が並層分裂することにより、向軸側にキールが形成される(A)。奇形葉では、窪みの内側に向かってキール様の突起ができることから、通常の葉で並層分裂を起こす仕組みが、異所的に袋の内側の表皮に近い細胞で起こることによって引き起こされたのかもしれない(B、C)。一方、キール様突起の先端が3つに分かれる理由はよくわからない。同じサラセニア科のヘリアンフォラ属は2枚のキールを形成するが(E)、襟の組織と繋がっており、奇形葉の先端部のみが3叉になるのとは異なっているように思われる。フクロユキノシタの捕虫葉の向軸側にできるキールは柄があり、先端が二叉になるが、中央の突起(C、Dの青色星印)は見当たらない。しかし、平面上の原基の二叉にする点では、似たような仕組みを使っている可能性もある。サラセニアの奇形葉はこれ以上研究することが難しいが、フクロユキノシタは無菌培養系も確立され、ゲノムも解読されているので(Fukushima et al. 2017)、二叉に分かれるキールの仕組みは植物の発生を考える上で新しい仕組みの解明につながるかもしれない。(A)、(B)は福島健児博士撮影。(H)はLloyd (1942)より改図。
In normal pitcher leaves, the keel is formed on the adaxial side by periclinal cell divisions (A). In the malformed leaf, a keel-like protrusion is formed toward the inner side of the depression, suggesting that the mechanism of periclinal cell division in normal leaves may be ectopically coopted in subepidermal tissue of the inner side of the pitcher (B, C). On the other hand, it is more unclear why the tip of the keel-like structure is divided into three parts. The genus Helianthus, also in the Sarraceniaceae, forms two keels (E), but they are divided from the connected point to the collar, which seems to be different from the trifurcation of only the apical part of the malformed leaf. The keel formed on the adaxial side of pitcher leaves of the Albany pitcher plant, Cephalotus follicularis is bifurcated at the tip (green arrow in [G, H] and a lateral keel that does not divide is shown in red arrow), but the central projection (blue star in C and D) is not visible. However, it is possible that they use a similar mechanism in terms of bifurcating the protuberance on the plane. While it is difficult to study the malformed leaves of Sarracenia further, it should be possible to study the mechanisms of bifurcated keel in Cephalotus follicularis, since a sterile culture system has been established and the genome has been available (Fukushima et al. 2017) and the mechanism of the bifurcated keel may lead to the elucidation of a new mechanism for plant development. Photos in (A) and (B) were taken by Dr. Kenji Fukushima. (H) is modified from Lloyd (1942)。
References:
Esau, K. (1977). Anatomy of Seed Plants (2nd ed.). John Wiley & Sons, Inc.
Fukushima, K., Fang, X., Alvarez-Ponce, D., Cai, H., Carretero-Paulet, L., Chen, C., Chang, T.-H., Farr, K. M., Fujita, T., Hiwatashi, Y., Hoshi, Y., Imai, T., Kasahara, M., Librado, P., Mao, L., Mori, H., Nishiyama, T., Nozawa, M., Pálfalvi, G., … Hasebe, M. (2017). Genome of the pitcher plant Cephalotus reveals genetic changes associated with carnivory. Nat. Ecol. Evol., 1: 59. http://dx.doi.org/10.1038/s41559-016-0059
Fukushima, K., Fujita, H., Yamaguchi, T., Kawaguchi, M., Tsukaya, H., & Hasebe, M. (2015). Oriented cell division shapes carnivorous pitcher leaves of Sarracenia purpurea. Nat. Commun., 6: 6450.
Fukushima, K., Narukawa, H., Palfalvi, G., & Hasebe, M. (2021). A discordance of seasonally covarying cues uncovers misregulated phenotypes in the heterophyllous pitcher plant Cephalotus follicularis. Proc. Royal Soc. B: Biol. Sci. 288: 20202568. https://doi.org/10.1098/rspb.2020.2568
Lloyd, F. E. (1942). The Carnivorous Plants. Dover Publications, Inc.
Whitewoods, C. D., Whitewoods, C. D., Gonçalves, B., Cheng, J., Cui, M., & Kennaway, R. (2020). Evolution of carnivorous traps from planar leaves through simple shifts in gene expression. Science 367: 91–96.
長谷部光泰. (2020). 陸上植物の形態と進化. 裳華房.
カテゴリー:未分類|サラセニアの奇形葉 Malformed leaf of Sarracenia はコメントを受け付けていません
同僚の先生から、「研究所の隣のホテルの前で咲いている、シソのような花を付けた裸子植物(A)はなんですか」と聞かれた。さっそく見に行くと、エリカ コロランスErica coloransのホワイトデライト“White Delight”という品種だった。エリカ コロランスは、シソ科や裸子植物とは縁遠く、ツツジ科ツツジ亜科エリカ連に属する(B: Gillespie and Kron 2010)。エリカ連の種は、花粉が粘って糸を引くことがない形質が共通しており(Stevens 2001 onwards on 10 Oct. 2023)、カルーナ属 Calluna(C、D)、ダボエキア属 Daboecia、エリカ属 Erica (A)の3属からなる(Stevens et al. 2004)。エリカ連の種はイギリスなどのヒース heathの主要構成樹種である(D:イギリス、Yorkshire Dales国立公園のマラム・コーヴ Malham Coveに群生するCalluna vulgaris)。
I was asked by a colleague about a gymnosperm. with flowers resembling those of the mint family (Lamiaceae) (A) , which was blooming in front of the hotel next to our research institute. I went to see it immediately and found out it was Ericacolorans “White Delight.” Ericacolorans is distant from the Lamiaceae or gymnosperms, and it belongs to the tribe Ericeae, the subfamily Ericoideae of the heath family Ericaceae (B: Gillespie and Kron 2010). Species within Ericeae share a common characteristic of not producing sticky and thread-drawing pollen (Stevens 2001 onwards on 10 Oct. 2023). This subfamily comprises three genera: Calluna (C, D), Daboecia, and Erica (A) (Stevens et al. 2004). Species within Ericeae are the main components of heath ecosystems in countries like the UK (D: Calluna vulgaris forming extensive colonies at Malham Cove in the Yorkshire Dales National Park in the UK).
エリカ連の3属は、花の数生、花冠が宿在するか、葉序、葉の基部が茎に沿着するかどうか(AのCalluna vulgarisの葉の基部は、黄色矢印部分で沿着しており、BのErica cristataは葉柄[白色矢印]が明確で葉身基部は沿着していない)、エリカ葉を形成するかで分類できる(Stevens et al. 2004)。
The three genera of Ericeae can be classified by the number of flowers, whether the corolla is scarious and persistent, the phyllotaxy, whether the leaves are petiolate or tailed (A, Calluna vulgaris: the sessile leaves are tailed along the stem at the yellow arrowhead; B, Erica cristata: the petiole [white arrow] is prominent and the leaf base is not tailed), and whether they form ericoid leaves (Stevens et al. 2004).
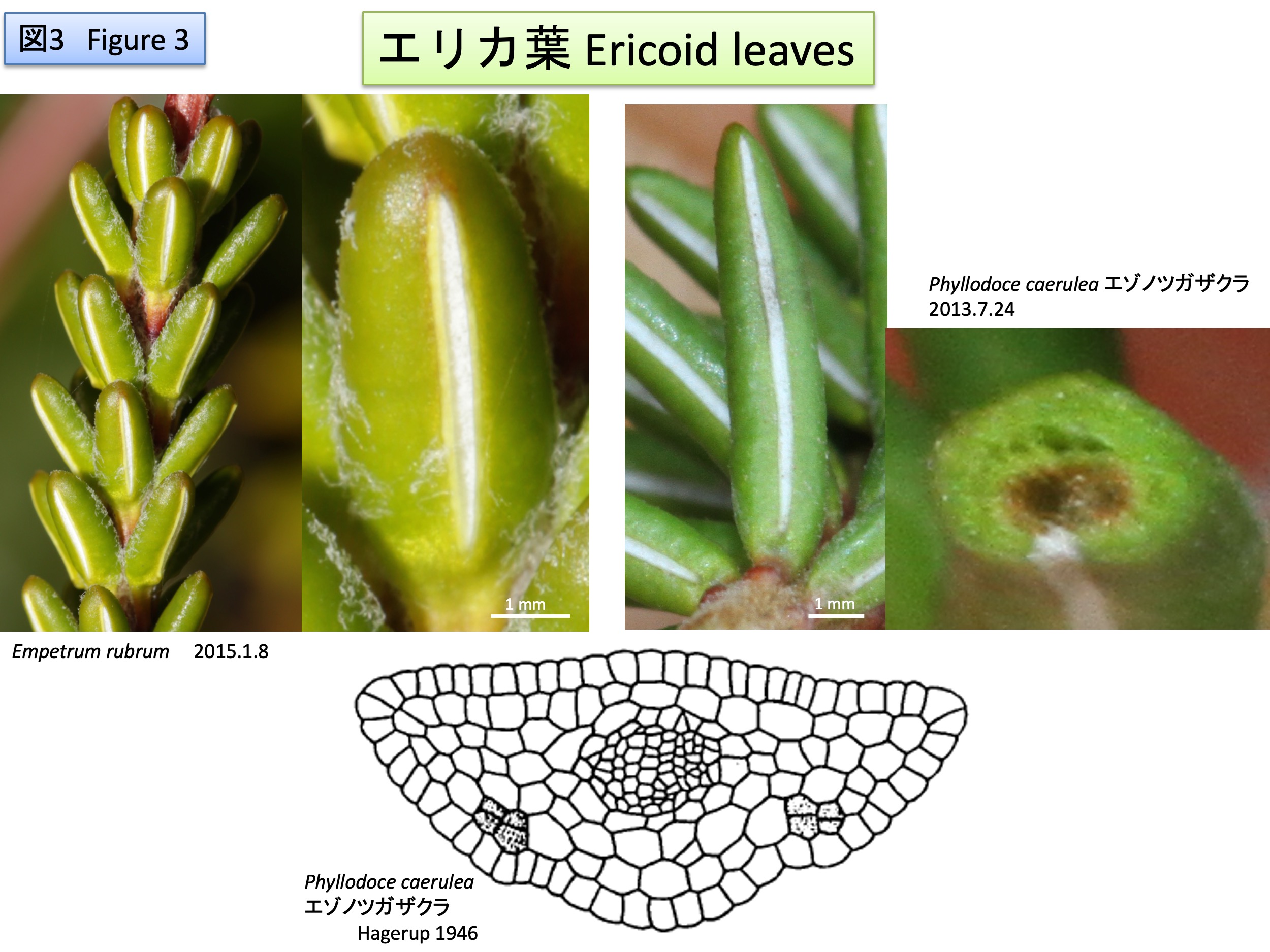
Species in Erica、Empetrum and Phyllodoce form leaves with enation. Protruded parts curved to the abaxial side to cover abaxial surface. These species are distributed in the area with strong wind and the morphology is beneficial to survive in the environment to keep moisture to the abaxial surface with stomata (Bocher 1981). Meristematic activity is changed from original leaf margins to abaxial regions shown in dots (Hagerup 1946).
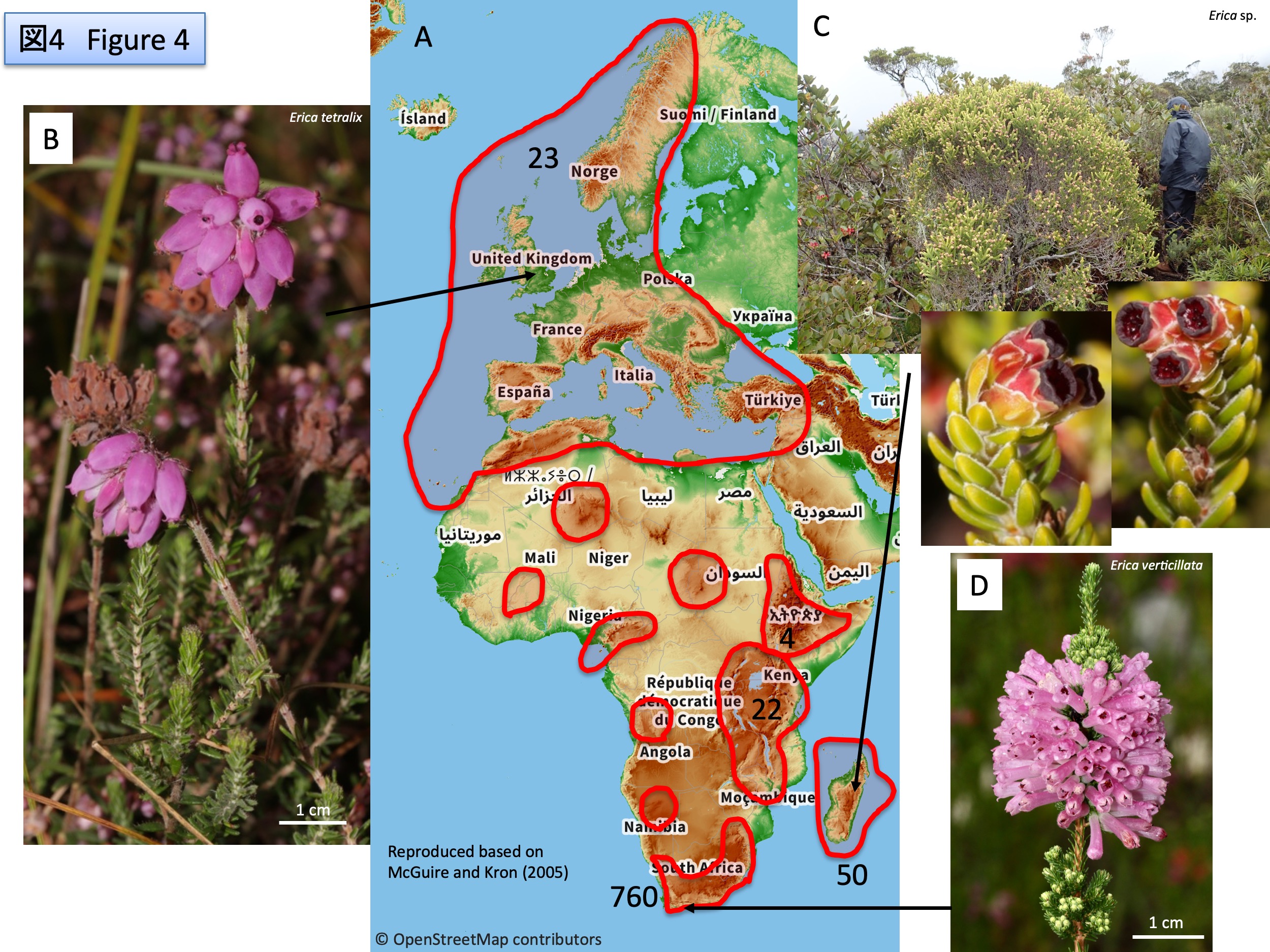
エリカ属 Ericaは、約865種からなり、北欧から、アフリカの熱帯地方の高山を経て、マダガスカル、南アフリカまで、南北に分布する(McGuire and Kron 2005)。南アフリカでは約760種に適応放散している。分布図(A)は、McGuire and Kron (2005)を改図し、種数の多い分布域に数字で種数を示した。(B)はイギリスのヨークシャー地方に自生するErica tetralix、(C)はマダガスカルのMarojeji山に自生し風媒花の種、(D)は南アフリカに自生するErica verticellata。
The genus Erica consists of ca. 865 species and has a north-south distribution from northern Europe, through the high mountains of tropical Africa, to Madagascar and South Africa (McGuire and Kron 2005). In South Africa, Erica has adapted and dispersed to about 760 species. Distribution map (A) is modified from McGuire and Kron (2005), with the number of species indicated by numbers in the distribution areas with the relatively higher number of species. (B) is Erica tetralix, native to Yorkshire, England; (C) is a wind-pollinated species native to Mount Marojeji, Madagascar; (D) is Erica verticellata, native to South Africa.
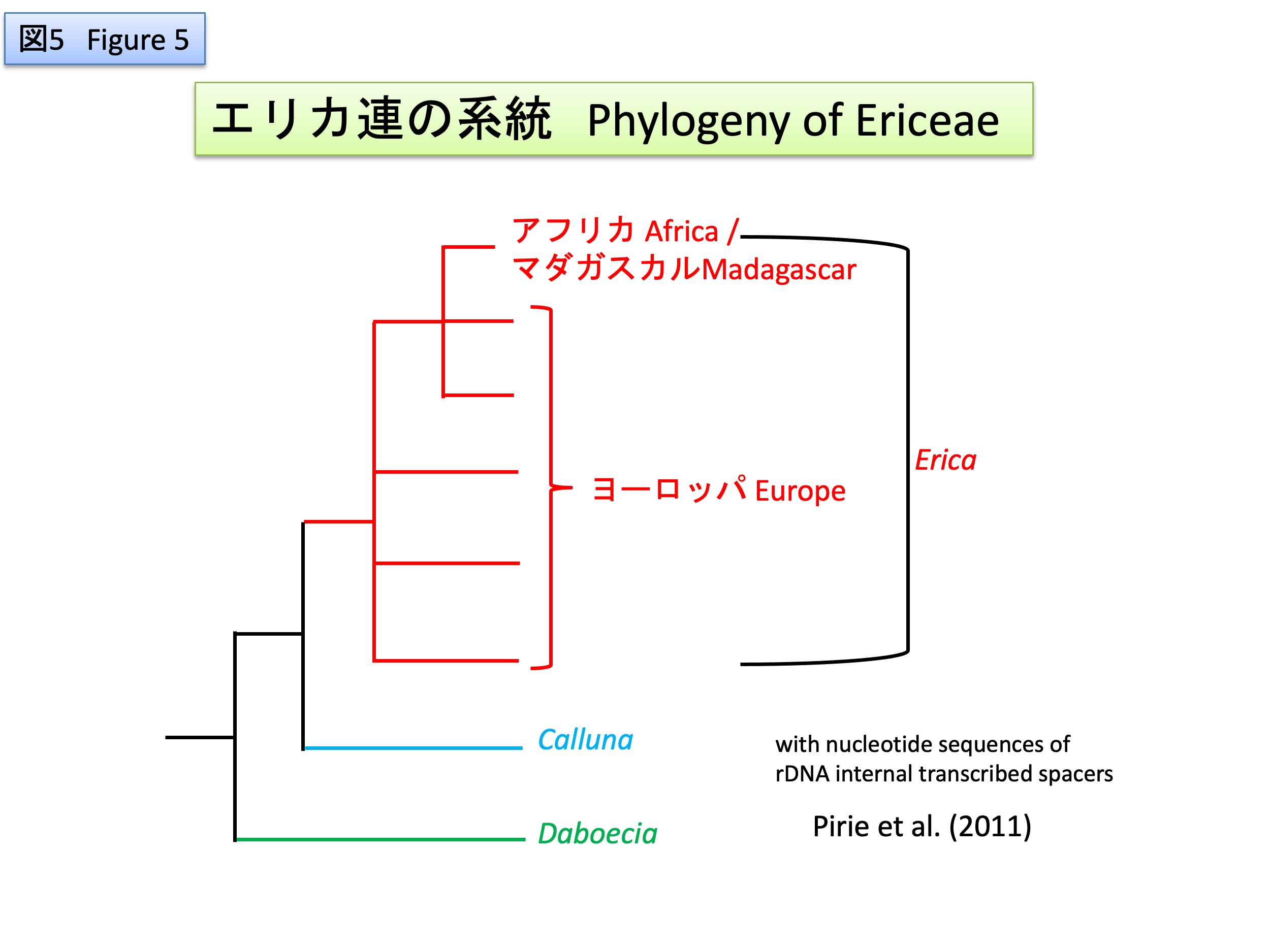
葉緑体DNAや核のrDNAの遺伝子間領域の塩基配列比較から、エリカ連の中で、デボエキア属が最初に分岐し、その後、カルーナ属とエリカ属が分岐したと推定されている(McGuire and Kron 2005, Pirie et al. 2011)。エリカ属の中では、ヨーロッパに分布する種が基部で側系統となり、アフリカとマダガスカルに分布する種類は系統樹の末端で単系統群となることから、エリカ属の共通祖先はヨーロッパに起源したと推定されている(McGuire and Kron 2005, Pirie et al. 2011)。また、西ヨーロッパの5600万年前の地層からエリカ属の花粉が産出する(Hofmann 2018)ことから、大陸移動により、チーシス海が閉じ、ヨーロッパとアフリカに陸橋のできた新生代中新世中期(約1700万年前)(Levyns 1964)にヨーロッパからアフリカへ分布を広げたのではないかと推定されている(McGuire and Kron 2005)。
Based on sequence comparisons of chloroplast genes and intergenic regions of nuclear rDNA, it has been inferred that the genus Deboecia was the first to diverge within the tribe Ericeae, followed by Caruna and Erica (McGuire and Kron 2005, Pirie et al. 2011). Within the genus Erica, the common ancestor of Erica is presumed to have originated in Europe, as the species distributed in Europe are lateral at the base, while the species distributed in Africa and Madagascar form a monophyletic group at the terminal of the phylogenetic tree (McGuire and Kron 2005, Pirie et al. 2011). In addition, fossil pollen of Erica was found from 56 Ma (million years ago) -old strata in Western Europe (Hofmann 2018), suggesting that the genus have spread from Europe to Africa during the middle Cenozoic Miocene (about 17 Ma) (McGuire and Kron 2005), when continental drift closed the Tyasis Sea and created a land bridge between Europe and Africa (Levyns 1964).
ヨーロッパの種は釣り鐘型の花(A)を形成し、虫媒の可能性が高いと考えられており(McGuire and Kron 2005)、アザミウマが主たる媒介昆虫ではないかと推定されている(Hagerup and Hagerup 1953)。一方、南アフリカでは花形態が多様化し、長い筒状から短い壺型までさまざまな形態の花を持つ種が進化している。この多様性は、花粉送粉動物との共進化によってもたらされた可能性が高い。昆虫媒だけでも、壺形(B)や球形(C)でアザミウマなどによって花粉媒介される種に加え、花冠が筒状に伸びて(D、E)花冠よりやや長い口吻を持つハエによって媒介される種、雄ずいが花冠先端から突出し(F、G)ハチによって媒介される種などが見られる(Rebelo et al. 1985, Oliver 1991, Pirie et al. 2011)。(A) イギリスヨークシャー地方で撮影したEricatetralix、(B) Erica articularis、(C) Erica pulchella、(D) Erica cristata、(E) Erica cristata、(F) Erica ericoides、(G) Erica equisetifolia。(B)から(G)は南アフリカのFernkloof Nature Reserveにて撮影。
European species form bell-shaped flowers (A) and are likely pollinated by insects (McGuire and Kron 2005), with thrips presumed to be the primary vector (Hagerup and Hagerup 1953). On the other hand, floral morphology has diversified in South Africa, with species having evolved a variety of flower morphologies ranging from long tubular to short vase-shaped. This morphological diversity should have coevolved with pollinators: flowers with urn-shaped (B) or spherical (C) corollas are pollinated by thrips; tubular corollas (D, E) are by flies with slightly longer proboscises than corollas; flowers with protruding stamens are by by bees(F、G) (Rebelo et al. 1985, Oliver 1991, Pirie et al. 2011). (A) Erica tetralix native to Yorkshire, England, (B) Erica articularis, (C) Erica pulchella, (D) Erica cristata, (E) Erica cristata, (F) Erica ericoides, (G) Erica equisetifolia. (B) to (G) were taken at Fernkloof Nature Reserve, South Africa.
昆虫媒花よりも花冠が伸び、頑丈になった鳥媒花の種も多く見られる。図1AのErica coloransや図2DのErica verticillataに加え、 Erica patersonia(A)、Erica viscariasubsp. longifolia (B)、Erica perspicua(C)、Erica sessiliflora (D)、Erica cruenta(E)、Erica plukenetii subsp.penicellata(F)、Erica cerinthoides(G)など少なくとも約70種が鳥媒だと考えられている(Rebelo et al. 1985)。ズズメ目タイヨウチョウ科のオナガゴシキタイヨウチョウNectarinia violaceaが主たる媒介鳥だと考えられている(Rebelo et al. 1985, Oliver 1991)。
There are also a number of bird pollinated species with more elongated and robust corollas than insect pollinated species. In addition to Erica colorans (Figure 1A) and Erica verticillata (Figure 2D), Erica patersonia (A), Erica viscaria subsp. longifolia (B), Erica perspicua (C), Erica sessiliflora (D), Erica cruenta (E), Erica plukenetii subsp. penicellata (F), Erica cerinthoides (G), and at least about 70 other species are considered to be bird pollinated (Rebelo et al. 1985). The sunbird Nectarinia violacea is thought to be the primary pollinator (Rebelo et al. 1985, Oliver 1991).
In addition, about 20 species have evolved to form wind-pollinated flowers (A-E). Compared to insect- and bird-pollinated flowers, petals are reduced and the stigmas are enlarged and protruding (B-D). (A-D) Erica axillaris and (E) Erica muscosa were photographed at Fernkloof Nature Reserve, South Africa. Flower morphology is known to convergently evolve into similar forms for each pollinator, a phenomenon known as the pollination syndrome. In the South African lineage, flowers pollinated by thrips, long proboscis flies, bees, and birds as well as wind have evolved several times in parallel (Pirie et al. 2011), which may provide good material for analyzing the evolutionary mechanism of the pollination syndrome if genes regulating flower morphology can be studied.
References:
Bocher TW. 1981. Evolutionary trends in Ericalean leaf structure. Biol Skr kgl danske Vidensk Selsk 23: 1–64.
Gillespie, E. and Kron, K. 2010. Molecular phylogenetic relationships and a revised classification of the subfamily Ericoideae (Ericaceae). Mol. Phyl. Evol. 56: 343-354.
Hagerup O. 1946. Studies on the Empetraceae. Kgldanske Vidensk Selsk biol Medd 20: 1–49.
Hagerup, E. and Hagerup, O. 1953. Thrips pollination of Erica tetralix. New Phytol. 52: 1-7.
Hofmann, C.C. 2018. Light and scanning electron microscopic investigations of pollen of Ericales (Ericaceae, Sapotaceae, Ebenaceae, Styracaceae and Theaceae) from five lower and mid-Eocene localities. Bot. J. Linnean Soc. 187: 550-578.
Levyns, M.R. 1964. Migrations and origin of the Cape Flora. Trans. R. Soc. S. Afr. 37: 85-107.
McGuire, A.F. and Kron, K.A. 2005. Phylogenetic relationships of European and African Ericas. Int. J. Plant Sci. 166: 311-318.
Oliver, E.G.H. 1991. The Ericoideae (Ericaceae) – a review. Contrib. Bolus Herb. 13: 158-208.
Stevens, P.F. 2001 onwards. Angiosperm Phylogeny Website. 10th Oct. 2023, http://www.mobot.org/MOBOT/research/APweb/. Stevens, P.F. with Luteyn, J., Oliver, E.G.H., Bell, T.L., Brown, E.A., Crowden, R.K., George, A.S., Jordan, G.J., Ladd, P., Lemson, K., McLean, C.B., Menadue, Y., Pate, J.S., Stace, H.M., and Weiller, C.M. 2004 Ericaceae. pp. 145-194. In K. Kubitzki ed. The Families and Genera of Vascular Plants. VI.
The Japanese stone pine Pinus pumila, known as “Haimatsu“ as a Japanese name grows on ridgelines (A) and mountain peaks (B) beyond the forest limit (A, B: both refer to the ridgeline from Mount Suisho-dake to Mount Akaushi-dake in the Northern Alps, Japan). The fertilized pine cones (C) mature in the following year (D).
The cones of the Japanese stone pine, Pinus pumila, referred to as “Haimatsu” as a Japanese name have slender stalks that can be easily detached from the branches but are not typically found on the forest floor. On the other hand, when in the rocky areas and hiking trails within the P. pumila zone, it is common to come across scattered three to five fallen cones (E). Unlike many other pine species, P. pumila seeds lack wings (C). You may find some ruptured seed coats with empty interiors. The number of seeds on the ground is often much lower than the total number of seeds inside the cones. Furthermore, P. pumila seedlings tend to grow in clusters (E, F: F is an enlargement of the central part of E). To explain this phenomenon, Hayashida (2003) conducted research on Mount Apoi in Hokkaido. The Eurasian nutcracker, Nucifragacaryocatactes (G: photographed at Mount Senjo in Nagano Prefecture) starts damaging the cones and consuming immature seeds from mid-July when the cones are still immature. As August approaches, mature cones turn brown, and their stalks wither, making them easier to pick. Nutcrackers select the brown ones, crack them open with their beaks, and eat the contents of some seeds. Meanwhile, they pick out the well-developed seeds from the remaining ones and store them in a sublingual pouch. They collect 15 to 78 seeds from 2 to 10 cones in one feeding session. During the breeding season in August, the crows transport the seeds to the subalpine forest floor, their breeding site, and store them in several locations at a depth of a few centimeters. Some of these stored seeds are forgotten and end up germinating. However, since the subalpine zone is lower than the natural habitat of P. pumila and covered with forest litter, most of the seedlings perish. On the other hand, by September, after the breeding season, the large-billed crows start burying the seeds they collected near the P. pumila‘s natural habitat. Here, the forgotten seeds can grow, and P. pumila seed reproduction is successful. It is believed that nutcrackers can transport seeds up to approximately 1 km, making their seed dispersal highly efficient. Although other animals such as rock ptarmigans Lagopus muta (H) and bears also eat P. pumila seeds, 96% of the seeds are transported by nutcrackers. This unique seed dispersal strategy is observed not only in P. pumila but also in other species such as Pinus albicaulis, Pinus flexilis, Pinus edulis in North America, Pinus cembra in Europe, Pinus sibirica in Siberia, and Pinus koraiensis in East Asia (references are available in Hayashida [2003]).
Hayashida, M. 2003. Seed dispersal of Japanese stone pine by the Eurasian Nutcracker. Ornithol. Sci. 2: 33-40.
ヤマトグサ属Theligonumは、4種が知られている。日本と中国東部にヤマトグサ Theligonum japonicum Okubo & Makino (Chen and Funston 2011)、台湾にT. formosanum (Ohwi) Ohwi & T.S. Liu (Chen and Funston 2011)、中国中西部(四川省、雲南省など)にT. macranthum Franchet (Chen and Funston 2011)、地中海沿岸にT. cynocrambe L. (https://www.gbif.org/species/2919295)、が分布する。
Four species of the genus Theligonum are known: Theligonum japonicum Okubo & Makino (Chen and Funston 2011) in Japan and eastern China, T. formosanum (Ohwi) Ohwi & T.S. Liu (Chen and Funston 2011) in Taiwan, T. macranthum Franchet (Chen and Funston 2011) in central and western China (Sichuan Prov, and Yunnan Prov.), T. macranthum Franchet (Chen and Funston 2011) in central and western China (Sichuan, Yunnan, etc.), T. cynocrambe L. (https://www.gbif.org/species/2919295) on the Mediterranean coast.
(A-C) T. japonicum: Flowering individuals (A), enlarged flowers (B), and non-flowering individuals (C).
The genus Theligonum belongs to the family Rubiaceae, subfamily Rubioideae, and section Theligoneae. The section Theligoneae includes only the genus Theligonum.
図3 ヤマトグサの栄養シュートと生殖シュート Vegetative and reproductive shoots of Theligonum japonicum
T. japonicum is monoecious, with the same individual forming male and female flowers (A, B). In the vegetative shoot, the leaves are opposite (C, D), as in other Rubiaceae species. On the other hand, in the flowering reproductive shoot, two male flowers are formed in the opposite position to the leaf (E, F). The male flowers are hypothesized to be formed in the axils of the degenerated and lost leaves.
図4 ヤマトグサの雄花 Male flowers of Theligonum japanicum
成熟した雄花の蕾の先端をそっと触ると、ほんの数秒で、1輪のみある花被が背側(外側)に反り返り(GからJ:5秒ほどで開いた)、20本程度の雄ずいが垂れ下がる(J)。雄ずい数は花によって変異があり(Rutishauser et al. 1998: 内貴 2017)、花被が4枚なのに、雄ずい数が4の倍数でない場合もある。アカネ科の他の種では、雄ずい数は種内で一定の場合がほとんどであるが、ヤマトグサでどうして雄ずい数が変異するのかはよくわかっていない(Rutishauser et al. 1998)。
When the tip of a mature male flower bud is gently touched, in just a few seconds the single whorl perianth turns abaxially (outward) (G to J: opened in about 5 seconds), and about 20 male stamens hang down (J). The number of stamens varies from flower to flower (Rutishauser et al. 1998: 内貴 2017), and the number of stamens may not be a multiple of 4, even though the flower has 4 perianths. In most other species of the family Rubiaceae, the number of stamens is constant within a species, but it is not well understood why the number of stamens varies in the genus Theligonum (Rutishauser et al. 1998).
The filaments of stamens (K, L; L is an enlargement of the black dotted area of K) is a wiggly array of odd cells (M) with ridges and longitudinal stripes, about 50 µm in diameter and thin. They are soft and mobile, but strong and hard to break. Thus, when the wind blows, the anthers wobble and effectively disperse pollen.
図6 ヤマトグサの雌しべ Female flower of Theligonum japonicum
花柱(QとRの赤色星印)は花被(Qの黒色星印、R)に覆われる部分のみで、乳頭状細胞を持つ柱頭が長く伸長する。子房は発生初期は2室だが、1つの胚珠のみが発達し(S、T)、最終的に1室となる(Rutishauser et al. 1998)。
ヤマトグサ属は、他の多くのアカネ科の種と比較して、(1)葉序が常に対生ではなく、栄養シュートは対生だが、花序は互生へと転換すること、(2)虫媒ではなく風媒anemophyllyであること、(3)雄花、雌花ともに、花被が2輪ではなく1輪であること、(4)雄花が葉腋ではなく、葉腋の反対側から生じること、(5)雄花の雄ずいが1輪の花被と同数ではなく、2から19本もあること、(5)花柱がほとんど無く、円柱状に伸びた柱頭が子房基部から側生すること、(6)子房が2室が癒合して1室となり1胚珠のみ形成されることなどの点が異なっている(Rutishauser et al. 1998)。ただ、どの形質もアカネ科の中で見られる形質であり、アカネ科に含めるのが妥当であると提唱されたが(Wunderlich 1971)、異論もあった(Praglowski 1973, Nowicke and Skvarla 1979)。しかし、詳細な形態比較(Rutishauser et al. 1998)、分子系統解析(Bremer 2009)から、アカネ科のアカネ連Rubieae、ケロギア属Kelloggia、プトリア連Putorieaeと単系統群であることがわかった。ハシカグサに葉の外観が似るが、花はもとより、托葉形態、葉の毛の生え方が異なっており、分子系統解析からも姉妹関係ではない(Bremer 2009, Neupane et al. 2015)。
Female flowers are formed in the leaf axils of the reproductive shoot (A, B, N-Q). (Q, R) The ovary is inferior, but the perianths and pistil are formed not at the tip of ovary but the lateral side.
The perianth is transparent, membranous, and trace-like. The petals are thinner than the sepals in other Rubiaceous species, leading to the hypothesis that the sepals are degenerated (Rutishauser et al. 1998) or that the petals are degenerated and only the sepals remain (内貴 2017), but which is valid is not known. Further research is needed to determine which genes are expressed, those working in the sepals or those working in the petals.
The style is the only part covered by the petiole, with a long elongated stigma with papillose cells. The ovary is biloculate in early development, but only one ovule develops (S, T) and eventually becomes unilocular (Rutishauser et al. 1998). Compared to many other species in the family Rubiaceae, the genus Theligonum has the following special characteristics: (1) the leaves are not always opposite, the vegetative shoot is opposite, but the inflorescence converts to alternate, (2) it is wind pollinated (anemophily) rather than insect pollinated, (3) both male and female flowers have a single perianth whorl, (4) the male flowers are produced on the opposite side of the leaf, and no subtending leaf, (5) the number of stamens are not equal to that of perianths (four perianths), but 2 to 19, and (6) The ovary is bilocular but changes to actually unilocular with a single ovule. (Rutishauser et al. 1998). However, all of the traits as evolutionary trends are found in other Rubiaceae (Rutishauser et al. 1998), and it was proposed that they should be included in the family (Wunderlich 1971), but there were other hypotheses (Praglowski 1973, Nowicke and Skvarla 1979). However, detailed morphological comparisons (Rutishauser et al. 1998) and molecular phylogenetic analysis (Bremer 2009) have shown that Theligonum forms a clade with Rubieae, Kelloggia, and Putorieae. Although the leaves are similar in appearance to those of Neanotis hirsuta in the same family, morphology of the flowers, leaf and leaf hair are different, and molecular phylogenetic analysis indicates that they are not sister taxa (Bremer 2009, Neupane et al. 2015).
図7 アカネ科で風媒花のCoprosma repens Wind pollinated flowers of Coprosma repens in the Rubiaceae
アカネ科では、ヤマトグサ連の種以外に、Anthospermeae連で風媒花が知られている。Anthospermeaeに属する種は、丁字着葯で、葯糸が長く、垂れ下がった雄ずいを形成し(Rutishauser et al. 1998)、風媒花であると考えられている(Puff 1982, Thureborn et al. 2019)。アカネ科でヤマトグサ属以外は虫媒であると考えられていることから(Puff 1982, Thureborn et al. 2019)、Anthospermeaeの共通祖先で虫媒から風媒への進化がおこったと推定できる。
In addition to the species of Theligonum, wind-pollinated flowers are known in the tribe Anthospermeae of the family Rubiaceae; species belonging to the Anthospermeae have dorsifix anthers, long filaments, and pendulous stamens (Rutishauser et al. 1998) and are considered to be wind-pollinated (Rutishauser et al. Puff 1982, Thureborn et al. 2019). Since all members of the Rubiaceae are considered to be insect-pollinated (Puff 1982, Thureborn et al. 2019), it can be inferred that wind-pollinated flowers evolved at the common ancestor of the Anthospermeae from ancestral insect-pollinated flowers.
Compared to Anthospermeae species, Theligonum species are unique in that they have evolved multiple traits that contribute to wind pollination. These include numerous stamens, long and strong filaments, long and slender anthers that split open soon after flowering, tepals that are recurved to expose the stamens, pistils with the short style and the elongated stigma, reducted pistil perianths, and reproductive shoots in which the nodes where the male flowers are formed have no leaves on the male flower side, facilitating the dispersal of pollen. It is hypothesized that supergene formation may have been involved in the evolution of complex traits (for a review, 藤原 2023). Studying what genes control the multiple traits involved in wind pollination in Theligonum, and how these genes are arranged in the genome, may advance new perspectives on the evolution of multiple adaptive traits. Each of these traits is also a trait found in other series of the Rubiaceae, and it is possible that the genetic system possessed by the common ancestor of the Rubiaceae evolved as exaptation.
References:
Antonelli, A. et al. (2021). Settling a family feud: a high-level phylogenomic framework for the Gentianales based on 353 nuclear genes and partial plastomes. Am J Bot 108: 1143–1165.
Bordbar, F., Mirtadzadini, M., and Razafimandimbison, S.G. (2022). Phylogenetic re-assessment of the delimitation of Plocama and its species relationships and limits (Rubiaceae, Putorieae): resurrection of the monospecific genus Aitchisonia and a description of trib. nov. Aitchisonieae. Plant Systematics and Evolution 308: 1–11.
Bremer, B. 2009. A review of molecular phylogenetic studies of Rubiaceae. Ann. Missouri Bot. Gard. 96: 4-26.
Chen, J. and Funston, A.M. (2011) Theligonum 346-347, Wu, Z. Y., P. H. Raven & D. Y. Hong, eds. 2011. Flora of China. Vol. 19 (Cucurbitaceae through Valerianaceae, with Annonaceae and Berberidaceae). Science Press, Beijing, and Missouri Botanical Garden Press, St. Louis.
Neupane, S., Dessein, S., Wikström, N., Lewis, P.O., Long, C., Bremer, B., and Motley, T.J. (2015). The Hedyotis–Oldenlandia complex (Rubiaceae: Spermacoceae) in Asia and the Pacific: Phylogeny revisited with new generic delimitations. Taxon 64: 299–322.
Nowicke, J.W. and Skvarla, J.J. (1979) Pollen morphology: the potential influence in higher order systematics. Ann. Missouri Bot. Gard. 66: 633-700.
Praglowski, J. (1973) The pollen morphology of the Theligonaceae with reference to taxonomy. Pollen & Spores 15: 385-396.
Puff, C. (1982). The delimitation of the tribe Anthospermeae and its affinities to the Paederieae (Rubiaceae). Botanical Journal of the Linnean Society 84: 355–377.
Razafimandimbison, S.G., Kainulainen, K., Senterre, B., Morel, C., and Rydin, C. (2020). Phylogenetic affinity of an enigmatic Rubiaceae from the Seychelles revealing a recent biogeographic link with Central Africa: gen. nov. Seychellea and trib. nov. Seychelleeae. Mol Phylogenet Evol 143: 106685.
Rutishauser, R., Ronse Decraene, L.P., Smets, E., and Mendoza-Heuer, I. (1998). Theligonum cynocrambe: Developmental morphology of a peculiar rubiaceous herb. Plant Syst. Evol. 210: 1–24.
Rydin, C., Wikström, N., and Bremer, B. (2017). Conflicting results from mitochondrial genomic data challenge current views of Rubiaceae phylogeny. Am J Bot 104: 1522–1532
Thureborn, O., Razafimandimbison, S.G., Wikström, N., Khodabandeh, A., and Rydin, C. (2019). Phylogeny of anthospermeae of the coffee family inferred using clock and nonclock models. Int J Plant Sci 180: 386–402.
Wunderlich, R. (1971) Die systematische Stellung von Theligonum. Oesterr. Bot. Z. 119: 329-394.
内貴章世(2017) アカネ科、日本の野生植物第4巻(大橋他編)、平凡社、pp. 266-293, PL. 206-228.
Aristolochia giganteaはブラジルのBahia州やMinas Gerais州に分布する(Costa and Hime 1981)。手のひらの倍以上もある大きな花を付ける。花弁の模様には多型があり、左上の写真のように網目が顕著な個体や、中央の写真のように網目がほとんど目立たない個体がある。
ウマノスズサの仲間は特殊な送粉様式を持っている(Oelschlägel et al. 2009)。花粉媒介昆虫は種によって異なるが、概ねハエの仲間である。くびれた花筒には、奥に向かった毛が密生している。他の花の花粉を付けたハエは、毛と同じ向きには動きやすいが、逆方向には、毛に逆らうことになって動きにくい。そのため、花の奥へ奥へと入り込む。ウマノスズクサの仲間は両性花だが、雌期と雄期に分かれている。開花直後は雌期である。花筒の奥へと入った虫は、内向きの毛のせいで、花の外へは出られない。雄ずいや雌ずいの周りには蜜腺があり、蜜を探してうろうろすると、体についた他個体の花粉が、成熟した雌ずいにくっつき、受粉がおこる。開花後、1日から数日で、雌期から雄期へと変化し、花粉が葯から放出される。花粉は、動き回るハエの体に付着する。この時、花筒の角度が大きくなったり、内向きに生えた毛がやわらかくなったり、しなびれたりする。すると、外へ出る障害がなくなり、ハエは花の外へと脱出し、別な花へと花粉を運ぶことになる。
A. giganteaの花弁、とりわけ、花筒の入り口部分(写真の白矢印)は肉のような色をしている。テンナンショウやラフレシアの仲間に見られるように、いかにも、肉が腐った匂いがただよい、ハエを誘っていそうな雰囲気である。同じウマノスズクサ属のA. grandifloraも大きな花を付け(右の写真)、花筒入り口の色が濃く、腐肉臭を発する(Burgess et al. 2004)。
なので、A. giganteaの花の匂いを初めて嗅いだときには驚いた。柑橘系の良い香りがするのだ。
香りが異なるとすると、ハエ以外の訪花昆虫を呼び寄せているのだろうか。しかし、ブラジルBahia州Chapada Diamantinaの自生地での調査によると、下水溝やゴミ溜めで繁殖するノミバエ科Phoridaeのメガセリア属Megaseliaの種が花の中に入り、花粉を媒介しているという (Hipólito et al. 2012)。
Aristolochia gigantea is found in the states of Bahia and Minas Gerais in Brazil (Costa and Hime 1981). A. gigantea produces large flowers that are more than twice the size of the palm of one’s hand. The color pattern of the petals is polymorphic, with some individuals having pronounced reticulation, as in the upper left photo, and others having almost no reticulation, as in the center photo.
Members of the horse chestnut family have a special pollination mode (Oelschlägel et al. 2009). Pollinators vary among species, but are generally flies. The constricted flower tube is densely covered with hairs towards the back. Flies with pollen from other flowers can move easily in the same direction as the hairs, but not in the opposite direction, as they would be going against the hairs. This is why they penetrate deeper into the flower. They are bisexual, but are divided into female and male phases. Immediately after flowering, the flowers are in the female phase. Once inside the flower tube, the insects are unable to exit the flower due to the inward-facing hairs. There are nectar glands around stamens and gynoecium, and when they wander around looking for nectar, pollen from other individuals on their bodies sticks to the mature pistil and pollination occurs. One to several days after flowering, the flower changes from the female to the male phase, and pollen is released from the anthers. The pollen adheres to the body of the fly as it moves around. At this time, the angle of the flower tube increases, and the inwardly growing hairs become soft and pliable. Then there is no obstacle to exit, and the fly escapes out of the flower and carries pollen to another flower.
The petals of A. gigantea, especially the entrance of the flower tube (white arrows in the photos), are flesh-colored. As is the case with A. gigantea and A. rafflesia, it smells like rotting flesh, attracting flies. A. grandiflora, a member of the same genus, also has large flowers (photos on the right) with a dark color at the entrance of the flower tube and a carrion smell (Burgess et al. 2004).
So I was surprised when I first smelled the flowers of A. gigantea. It has a nice citrus scent.
If the scent is different, does it attract other flower-visiting insects besides flies? However, according to a study conducted in the native habitat of Chapada Diamantina in Bahia, Brazil, species of Megaselia species of the family Phoridae, which breed in sewage ditches and garbage dumps, enter the flowers and transmit pollen (Hipólito et al. 2012).
自生地では、A. giganteaの花は朝6時半ころ開花、その後、8時ころまでノミバエ類が多く訪花する(Hipólito et al. 2012)。他のウマノスズクサ類と同じように、開花初日は雌期で、花筒から入り込んだノミバエは、花筒の奥の雌しべの周辺でうろうろし、蜜腺から分泌される蜜を吸いながら、他個体の花粉を雌しべに受粉させる。翌日、葯が裂開して雄期になると、蜜腺から蜜が分泌されなくなり、香気が著しく減少、ノミバエの仲間は体に花粉を付けて、外界へと出ていく(Hipólito et al. 2012)。
In its native habitat, A. gigantea flowers around 6:30 in the morning, after which many flea flies visit the flowers until around 8:00 (Hipólito et al. 2012). As in other horse chestnut species, the first day of flowering is the female stage, when fruit flies enter through the flower tube and hang around the pistil at the back of the tube, sucking nectar from the glands and pollinating the pistil with pollen from other flowers. The next day, when the anthers split open and enter the male stage, the nectar glands no longer secrete nectar, the fragrance is significantly reduced, and the flea flies attach pollen to their bodies and leave for the outside world (Hipólito et al. 2012).
A. giganteaの花筒にも、内側に向かった毛が密生している(開花初日の写真)。しかし、自生地での調査によると、雌期でもノミバエ類は自由に出入りしており、雄期になっても花筒の毛はしなびれないという(Hipólito et al. 2012)。ただ、栽培株を観察すると、開花初日にあった毛が、3日後にはしなびれていたので、種内多型があるのかもしれない(上の写真)。栽培株の場合も、毛は花筒を完全に塞ぐほどは長くなく、ノミバエも無理すれば脱出できるようにも見える。従って、毛によって絶対的に捕らえているというわけではなく、外へ出にくくしつつ、蜜を与えることで、雄期になるまでノミバエを花の中に滞在させているのだろう。
The corolla tube of A. gigantea is densely covered with inward-facing hairs. However, according to a survey in the native habitat, flea flies freely move in and out even in the female stage, and the hairs on the flower tube do not become shriveled even in the male stage (Hipólito et al. 2012). However, when I observed the cultivated plants, the hairs that were present on the first day of flowering were shriveled three days later, so there may be intraspecific polymorphism (see photos above). In the case of the cultivated plants, the hairs were not long enough to completely block the flower tube, and the flea flies seemed to be able to escape if they were forced to (Hipólito et al. 2012). Therefore, the hairs are not absolutely trapping the fruit fly, but rather keeping it inside the flower until it reaches the male stage by providing nectar and making it difficult for it to escape.
ウマノスズクサ科は種類によって、訪花する昆虫に雌雄差がある。Aristolochia giganteaの場合、96%は雄である(Martin et al. 2017)。雌の出す誘引フェロモンに似た化学物質を出している可能性もあるかと思ったが、解析例は見つからなかった。
さらに奇妙なことに、柑橘類の匂いは、花弁の肉のような色をした組織から、腐臭はレモン色をした花筒入り口や内部の組織から放出されており、見た目と香りが一致しないという(Martin et al. 2017)。なんとも不可解な花である。
Depending on the species, the sex of pollinators are varied in Aristolochia, and in the case of Aristolochia gigantea, 96% are male (Martin et al. 2017). I wondered if they might be producing chemicals similar to the attractant pheromones produced by females, but I could not find any references.
Now, shall we consider why does A. gigantea attract flies when it smells fresh and citrusy instead of carrion?
A. gorgona, native to Costa Rica and Panama, was confused with A. gigantea for a long time, but in 2002 it was described as a separate species because of its larger flowers and the way the lower part of the corolla bends backward like a reflex when in bud (Blanco 2002). Although the species are so similar, the flowers of A. gorgona are said to have a putrid odor, unlike A. gigantea (Blanco 2002).
In 2017, the scent components of A. gigantea were revealed. Indeed, the main components of the aroma were (E)-citral, (Z)-citral, citronellol, and citronellal, which smell like sweet citrus fruits (Martin et al. 2017). However, dimethyl disulfide, a garlic-like odor found in rotting meat and rotten fruit, and 2-heptanone, the primary scent of Gorgonzola, were also detected, albeit in trace amounts, and these may contribute to attracting flea flies.
So why are so many citrus smells being released? There are several hypotheses, such as that the rotting odor is more effective when mixed with citrus scents, or that the native fruit flies prefer citrus colors and smells, but we do not know the answer.
Even stranger, the citrus smell is emitted from the flesh-colored tissue of the petals, while the rotten smell is emitted from the lemon-colored tissue at the entrance and inside of the flower tube, and the appearance and scent do not match (Martin et al. 2017). It is a very puzzling flower.
A. giganteaの花冠の模様は、アントシアニンと推定される色素が蓄積される細胞とされない細胞ができることによって形成されている。色素の蓄積は葉脈と葉脈の周辺でおこっている。キンギョソウの花弁で葉脈に沿ってアントシアニンが合成され、模様が入る仕組みが報告されている。bHLH転写因子DELILAとMUTABILISが表皮細胞で発現しているが、葉脈から表皮に向かって放射状発現するMYB転写因子VENOSAの発現と重なる部分でアントシアニン合成酵素を誘導すると考えられている (Shang et al., 2011)。A. giganteaの花冠の模様の多型の原因は、どの葉脈でどのくらい遠くまで色素合成が広がっているかによっている。キンギョウソウと同じ仕組みでできているのだとすると、VENOSAに対応する葉脈から表皮へシグナルを送る因子の広がり方によって多型が生み出されている可能性がある。VENOSAの発現がどのように制御されているか、すなわち、葉脈から表皮へのシグナルがどのような因子によってどのように担われているかは発生学的にも興味深いがまだわかっていない。
The coloration of A. gigantea is formed by the accumulation of pigments, presumably anthocyanins, in some epidermal cells and not in others. The pigment accumulates on and around the leaf veins. In snapdragons, the bHLH transcription factors DELILA and MUTABILIS are expressed in epidermal cells, but they are expressed at the same time as the MYB transcription factor VENOSA, which is expressed radially from the leaf veins to the epidermis (Shang et al., 2011). The cause of the polymorphism in the corolla pattern of A. gigantea depends on which veins and how far pigment synthesis is spread. If the mechanism is the same in A. gigantea as in snapdragons, the polymorphism may be produced by the spread of factors that signal from the veins to the epidermis in response to VENOSA. How the expression of VENOSA is regulated, i.e. by which factors the signal from the veins to the epidermis is carried, is of great developmental interest but is not yet known.
好奇心を触発していただき、材料を提供いただいた樋渡琢真博士に心より感謝申し上げます。I deeply appreciate Dr. Takuma Hiwatashi for inspiring my curiosity and providing living materials.
Blanco, M.A. (2002). Aristolochia gorgona (Aristolochiaceae), a new species with giant flowers from Costa Rica and Panama. Brittonia 54, 30–39.
Burgess, K., Singfield, J., Meléndez, V., and Kevan, P. (2004). Pollination biology of Aristolochia grandiflora (Aristolochiaceae) in Veracruz, Mexico. Ann. Missouri Bot. Gard. 91: 346-356.
de Costa, E.L., and da Hime, N.C. (1981). Biologia floral de Aristolochia gigantea Mart. et Zucc. (Aristolochiaceae) I. Rodriguesia 33, 23–69.
Hipólito, J., Viana, B.F., Selbach-Schnadelbach, A., Galetto, L., and Kevan, P.G. (2012). Pollination biology and genetic variability of a giant perfumed flower (Aristolochia gigantea Mart. and Zucc., Aristolochiaceae) visited mainly by small diptera. Botany 90, 815–829.
Martin, K.R., Moré, M., Hipólito, J., Charlemagne, S., Schlumpberger, B.O., and Raguso, R.A. (2017). Spatial and temporal variation in volatile composition suggests olfactory division of labor within the trap flowers of Aristolochia gigantea. Flora 232, 153–168.
Oelschlägel, B., Gorb, S., Wanke, S., and Neinhuis, C. (2009). Structure and biomechanics of trapping flower trichomes and their role in the pollination biology of Aristolochia plants (Aristolochiaceae). The New Phytologist 184, 988–1002.
Shang, Y., Venail, J., Mackay, S., Bailey, P.C., Schwinn, K.E., Jameson, P.E., Martin, C.R., and Davies, K.M. (2011). The molecular basis for venation patterning of pigmentation and its effect on pollinator attraction in flowers of Antirrhinum. The New Phytologist 189, 602–615.
ツチトリモチ科は世界の亜熱帯から熱帯にかけて分布し、13属、約42種が知られる(Kuijit and Hansen 2014, Nickrent 2020)。従来、ツチトリモチ科に含まれていたDactylanthus, Hachettea, Mystropetalonは他のツチトリモチ科の属とは異なった単系統群であることがわかり、Mystropetalaceae ミストロペタラ科を形成する(Su et al. 2015, Nickrent 2020)。
The Balanophoraceae includes ca. 42 species in 16 genera mainly in subtropical to tropical areas all over the world (Kuijit and Hansen 2014, Nickrent 2020). Dactylanthus, Hachettea, Mystropetalon, previously included in the Balanophoraceae were found to form a monophyletic group: the Mystropetalaceae (Su et al. 2015, Nickrent 2020).
Kuijt, J. and Hansen, B. 2014. Balanophoraceae. 193-208. In Kuijt, J. and Hansen, B. (eds). Series ed. K. Kubitzki, The Families and Genera of Vascular Plants. XII. Santalales, Balanophorales. Springer.
Nickrent, D.L. (2020). Parasitic angiosperms: How often and how many? Taxon 69, 5–27.
Su, H.J., Hu, J.M., Anderson, F.E., Der, J.P., and Nickrent, D.L. (2015). Phylogenetic relationships of Santalales with insights into the origins of holoparasitic Balanophoraceae. Taxon 64, 491–506.
Species in the Balanophoraceae induce host root growth inside of the parasitic rhizome, while other parasitic plants penetrate their tissue into the host (Hsiao 1993、邑田 2017).
Hsiao, S.-C., Mauseth, J.D., and Gomez, L.D. (1993). Growth and Anatomy of the Vegetative Body of the Parasitic Angiosperm Helosis cayennensis (Balanophoraceae). Bull. Torrey Bot. Club 120, 295–309.
邑田仁 2017 ツチトリモチ科 大橋広好他編「日本の野生植物 4」平凡社 pp. 72-74.
ツチトリモチ属はアジアからアフリカ、オーストラリアに分布するが、アメリカ大陸には分布せず、約20種が知られる(Hansen 1972, Su et al. 2012, 邑田 2017)。
The genus Balanophora is mainly distributed in the temperate and tropical areas from Asia to Africa and Australia but not in America and about 20 species are recognized (Hansen 1972, Su et al. 2012, 邑田 2017).
Reproductive systems in the Balanophoraceae are variable. The family includes dioecious, androdioecious, and agamospermic species (Stebbins 1941, Hansen 1972, Kuwada 1928). This species distributed in Malesia is dioecious. This is an individual with a female inflorescence.
Hansen, B. (1972). The genus Balanophora J. R. & G. Forster a taxonomic monograph. Dansk Botanisk Arkiv 28, 1–188.
Kuwada, Y. (1928). An occurrence of restitution nuclei in the formation of embryo sacs in Balanophora japonica. Bot. Mag. Tokyo 42, 117–129.
邑田仁 (2017). ツチトリモチ科 大橋他編「日本の野生植物 第4巻」 pp. 72-74. 平凡社
Stebbins, G.L. (2014). Apomixis in the angiosperms Bot. Rev. 7, 507-542.
Su, H.J., Murata, J., and Hu, J.M. (2012). Morphology and phylogenetics of two holoparasitic plants, Balanophora japonica and Balanophora yakushimensis (Balanophoraceae), and their hosts in Taiwan and Japan. J. Plant Res. 125, 317–326.
雌花序は多数の雌花からなる。
The female inflorescence contains a lot of flowers.
雄株の塊茎。
An unopened male tuber including a male inflorescence.
塊茎の断面。中に雄花を付けた雄花序が入っている。
Sections of a male tuber with a male inflorescence with several male flowers.
Even in the species to form both male and female flowers, the distribution of each sexual flower is variable. In Balanophora fungosa, there are plants with both female and male flowers (H in the above picture) and those with only male flowers (M in the above picture) are found.
In Balanophora tobiracola, male flowers are dispersed in an inflorescence. The mechanisms to induce the male and female flowers and to form the different patterns are unknown.
ヤクシマツチトリモチ Balanophora yakushimensisの種子はキノコを食べるネズミなどによって散布されるのではないかと考えられてきた(Conran and Li 2012)。しかし、匂いが無く夜行性の齧歯類を誘引せず、赤い色によってシロハラTurdus pallidusやルリビタキTarsiger cyanurusを誘引し種子散布している(Suetsugu 2020)。リュウキュウツチトリモチは赤と白の個体があるが、それぞれどのように種子散布されているかはわかっていない。
Conran, J.G. and Li, J. (2012). A Preliminary Report of Small Mammal Frugivory on Balanophora harlandii (Balanophoraceae). Plant Divers. Resour. 34, 466–470.
Suetsugu, K. (2020). A specialized avian seed dispersal system in a dry-fruited nonphotosynthetic plant, Balanophora yakushimensis. Ecology 101, e03129.
リュウキュウツチトリモチの花は自家受精が可能で、アリやゴキブリによって自家受粉されている(Kawakita and Kato 2007)。また、ハエやメイガの仲間が他花受粉に貢献しており、特に、メイガは卵を産み付け腐敗したリュウキュウツチトリモチを餌として幼虫が育つ(Kawakita and Kato 2007)。
Kawakita, A., and Kato, M. (2002). Floral biology and unique pollination system of root holoparasites, Balanophora kuroiwai and B. tobiracola (Balanphoraceae). Am. J. Bot. 89, 1164–1170.
ビャクダン目の科の検索表(Kuijt and Hansen 2014を改編)Key to families
Kuijt, J. and Hansen, B. 2014. Balanophoraceae. 193-208. In Kuijt, J. and Hansen, B. (eds). Series ed. K. Kubitzki, The Families and Genera of Vascular Plants. XII. Santalales, Balanophorales. Springer.
Nickrent, D.L. (2020). Parasitic angiosperms: How often and how many? Taxon 69, 5–27.
基礎生物学研究所生物進化研究部門の長谷部光泰が作っている「植物発生進化学」の講義録です。Lecture notes of “Plant Development and Evolution” by Mitsuyasu Hasebe at National Institute for Basic Biology (NIBB), Japan.
間違いがありましたら、mhasebe (at mark) nibb.ac.jpまでご連絡いただければ幸いです。If you find any mistakes, kindly inform me to mhasebe (at mark) nibb.ac.jp
































 A. giganteaの花筒にも、内側に向かった毛が密生している(開花初日の写真)。しかし、自生地での調査によると、雌期でもノミバエ類は自由に出入りしており、雄期になっても花筒の毛はしなびれないという(Hipólito et al. 2012)。ただ、栽培株を観察すると、開花初日にあった毛が、3日後にはしなびれていたので、種内多型があるのかもしれない(上の写真)。栽培株の場合も、毛は花筒を完全に塞ぐほどは長くなく、ノミバエも無理すれば脱出できるようにも見える。従って、毛によって絶対的に捕らえているというわけではなく、外へ出にくくしつつ、蜜を与えることで、雄期になるまでノミバエを花の中に滞在させているのだろう。
A. giganteaの花筒にも、内側に向かった毛が密生している(開花初日の写真)。しかし、自生地での調査によると、雌期でもノミバエ類は自由に出入りしており、雄期になっても花筒の毛はしなびれないという(Hipólito et al. 2012)。ただ、栽培株を観察すると、開花初日にあった毛が、3日後にはしなびれていたので、種内多型があるのかもしれない(上の写真)。栽培株の場合も、毛は花筒を完全に塞ぐほどは長くなく、ノミバエも無理すれば脱出できるようにも見える。従って、毛によって絶対的に捕らえているというわけではなく、外へ出にくくしつつ、蜜を与えることで、雄期になるまでノミバエを花の中に滞在させているのだろう。



















