基礎生物学研究所




トランスポゾンとゲノムの再編成

自然栽培条件下で DNA トランスポゾン nDart1 が転移するタギング系統から選抜されたイネの Lgg 変異体は、顕性(優性)の大粒変異である。ほ場において栽培し結実した穂(A)と玄米(B)。
ゲノム中には多くの転移因子(トランスポゾン)が存在しているが、その多くは転移する事ができない。しかし稀にゲノムによる抑制機構をすり抜けて転移できるトランスポゾンが存在する。どのようにゲノムはトランスポゾンの制御しているのか、また転移によって引き起こされるゲノムの再編成は生物にどんな影響を与えているのかを調べている。さらに内在性トランスポゾンを用いてイネの遺伝子破壊系統を作出して、機能ゲノム学的解析も試みている。
多くの生物のゲノム中には多くのトランスポゾンが存在している。例えばヒトではおよそ 45%、イネでは 35% がトランスポゾン様の配列である。トランポゾンによるゲノムの再編成は、進化の原動力一つとなっていると考えられるが、トランスポゾンの転移は、ホストのゲノムにとって有害になるので、転移する能力はジェネティックやエピジェネティクに抑制されており、通常の成育条件下で転移する事はまれである(文献 9)。そこで転移できる DNA トランスポゾンに注目して、トランスポゾンによるゲノムのダイナミズムと遺伝子発現の制御機構の解明を明らかにすることを試みている。
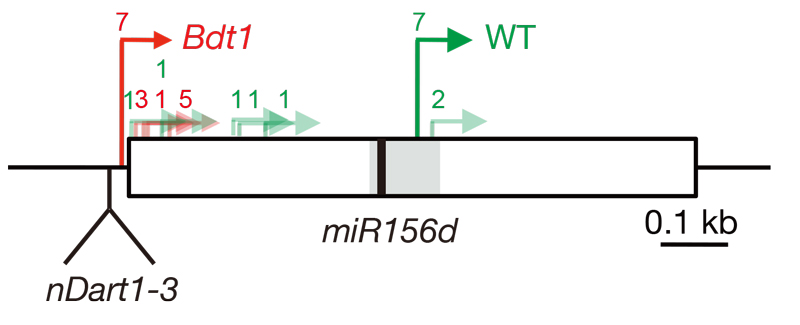
図 1. nDart の挿入による優性変異の原因の解明(文献 4)
高い精度でゲノム配列が決定されているイネは、トランスポゾンの挿入領域やゲノムの再編成を詳細に解析することができる。我々は自然栽培条件下で活発に転移することができる DNA トランスポゾン nDart1 を同定した。nDart1 の転移には、自律性因子 nDart1 が必要であるが、通常はエピジェネテイックに抑制されている。nDart1 が活発に転移する時期を明らかにし(文献 7)、さらに、脱メチル化によって nDart1 を持たないイネ系統でも転移を活性化できることも示した。nDart1 は、GC 含量の差が大きい領域に挿入し易い性質をもっているので、ゲノム中に存在している転移の制御因子の同定に向けて研究を行っている。
ゲノムの変異の多くは劣性となるが、nDart1 の挿入変異体の中にはしばしば優性となる突然変異体が観察される。不完全優性でわい性となる Bdt1 変異体では機能のあるマイクロ RNA の発現様式が nDart1 の挿入で変化していた(図1,文献6)。DNA トランスポゾンが優性変異の原因となる例は非常に珍しく、その原因は未解明な部分が残されているので、優性となった変異体を選抜して解析を行っている。
hATファミリーは、植物で発見され動物にまで存在している起源の古いDNAトランスポゾンです。イネ遺伝子の逆遺伝各的解析や活性なイネのDNAトランスポゾンを利用した共同利用研究に応じる事ができます。
栂根 一夫 助教 E-mail: tsugane@nibb.ac.jp TEL: 0564-55-7521

