研究の概要
植物が重力に応答する仕組みの解明
芽生えた場所で一生を過ごす植物は、重力、光、水分勾配などの外的環境を認識し、より効率的にリソースを獲得出来るように成長方向を調節しています。このような植物の応答は屈性と呼ばれています。本研究部門ではシロイヌナズナの重力屈性について、遺伝学、細胞生物学、分子生物学など様々な角度から研究を行なっています。細胞が重力方向をどのように認識し、生化学的情報に変換するか、またその情報をどの様に細胞から器官全体に伝達するかなど、植物の巧妙な重力方向の認識と成長制御のメカニズムを理解することを目指しています。
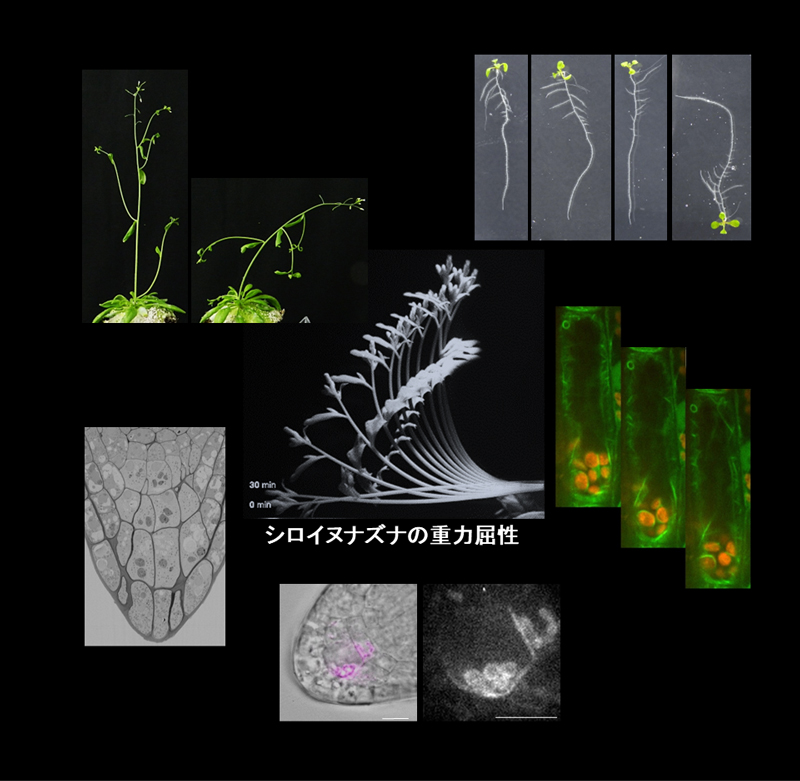
シロイヌナズナの重力屈曲と重力屈曲変異体。重力方向の認識に平衡石として働くと考えられているアミロプラスト。
植物の重力屈性とは
植物は重力方向を認識して、地上部は上向きに、根は下向きに成長する。重力方向は重力感受細胞内に存在するデンプン粒を蓄積した高比重のオルガネラであるアミロプラストが重力方向に移動することで感受される。その情報は細胞内シグナル伝達過程(重力シグナリング)を経て、植物ホルモンであるオーキシンの方向性を持った細胞間輸送の制御へと変換されると考えられている。私たちは重力感受と重力シグナリングに注目して、分子生物学的解析を初めとした多角的なアプローチにより重力屈性の分子機構の解明を目指している。
アミロプラストの沈降に伴う細胞内動態
重力方向を感受する細胞である花茎の内皮細胞や根端のコルメラ細胞において、アミロプラストの重力方向への移動には、液胞膜や細胞骨格の動態が適切に制御されることが重要である。私たちは、垂直ステージ共焦点顕微鏡を独自に構築し、重力方向の変化に伴う重力感受細胞内のオルガネラやタンパク質動態を詳細に観察することで、重力感受機構の理解を進めている(図1)。

図1.垂直ステージ共焦点レーザー顕微鏡で観察した内皮細胞
顕微鏡全体が90度回転しているため、成長時の重力方向を維持したまま細胞内を観察することができる(左図)。アミロプラスト(赤)が茎の重力感受細胞で重力方向に沈んでいる様子(右図)。GFPを用いて、液胞膜(左側)とアクチン繊維(右側)を可視化した。
重力シグナリングの分子機構
重力感受細胞に着目したトランスクリプトーム解析から、花茎、胚軸、根における重力シグナリングに関与するLZY遺伝子ファミリーを私たちは同定し、根や側枝の成長方向の決定はこの遺伝子ファミリーの制御下にあることを明らかにしてきた。最近、LZY蛋白質は根端にある重力感受細胞ではアミロプラストと細胞膜に見出され、アミロプラストから細胞膜へと移動する様子を私たちは捉えた(図2A)。このことは、アミロプラストが重力方向へ移動した先にある細胞膜、つまり重力方向側の細胞膜にLZY蛋白質が供給されることを意味する。アミロプラスト沈降という変化が重力感受細胞においてどのように認識されるのかは100年来の謎であったが、この発見と、オーキシン輸送に関連するRLD蛋白質を細胞膜上に呼び込むLZYの機能の発見により、重力に応答したオーキシンの極性輸送の仕組みが遂に分子的に説明可能となった(図2B)。現在、RLDのオーキシン輸送に関する分子機能の解明や、地上部における重力シグナル伝達も根と同様の仕組みにより行われているかについて検証を進めている。
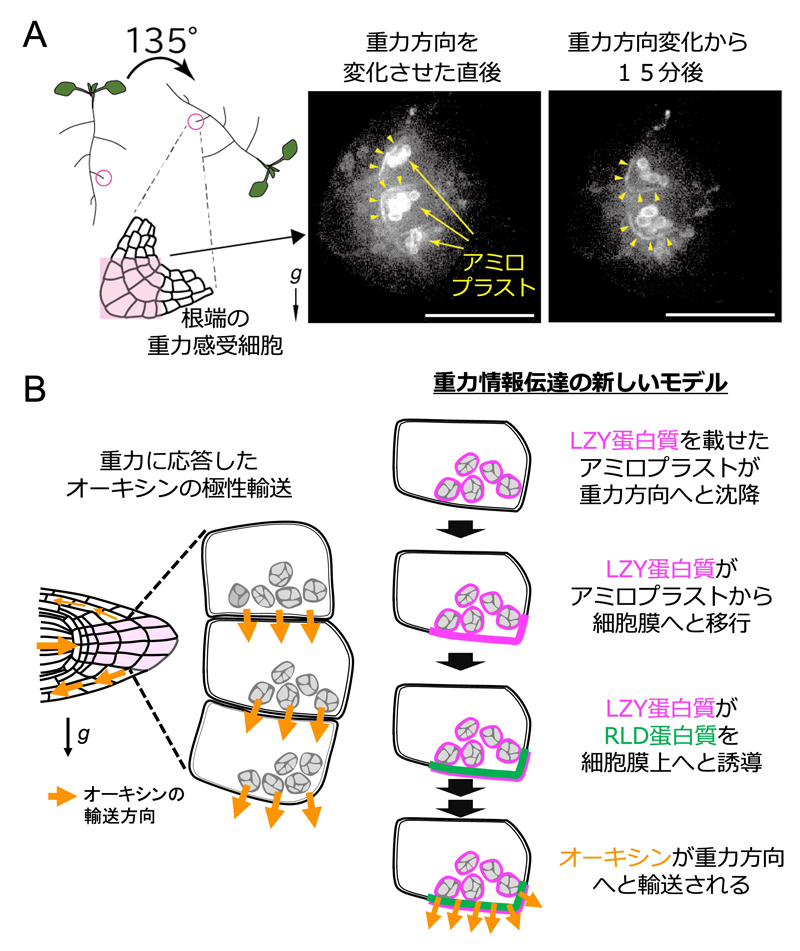
図2.重力方向の変化に伴う根の重力感受細胞におけるLZY蛋白質の細胞内局在の変化と重力情報伝達に関する新しいモデル
(A)蛍光蛋白質で標識したLZY蛋白質を発現する植物体を135度回転させ、重力方向を変化させた直後、15分後に根の重力感受細胞を蛍光観察した像。黒矢印が重力方向。黄矢印と黄矢尻は、アミロプラスト或いは細胞膜に存在するLZY蛋白質をそれぞれ示す。重力方向変化直後は元の重力方向側にアミロプラストが位置し、アミロプラスト及びその周辺の細胞膜上にLZY蛋白質が見出される。15分後には新たな重力方向にアミロプラストが沈降し、その周辺の細胞膜上にLZY蛋白質が見出される。 (B)明らかになった植物の重力情報伝達経路。
重力屈性と抗重力屈性
LZYは、根の下方向への成長、地上部(花茎)の上方向への成長を促進する働きをもつ。興味深いことに、
lzy多重変異体では野生型と逆に、根は上方向、地上部は下方向へと成長する傾向にある。植物の形態は、重力屈性およびそれとは逆方向へ成長させる仕組み(抗重力屈性)のバランスにより維持されていると近年考えられている。私たちは、上述の
lzy多重変異体が示す振る舞いは抗重力屈性がより顕著に現れた結果と捉え、その解析を通じて抗重力屈性の機構解明に取り組んでいる。
参考文献
西村岳志、四方明格、森田(寺尾)美代(2024)植物はどのように重力方向を感知するのか?-デンプン平衡石による植物の重力感知の仕組み「科学」岩波書店94, 152-157.
Nishimura, T., Mori, S., Shikata, H., Nakamura, M., Hashiguchi, Y., Abe, Y., Hagihara, T., Yoshikawa, H. Y., Toyota, M., Higaki, T., and Morita, M. T. (2023). Cell polarity linked to gravity sensing is generated by LZY translocation from statolith to the plasma membrane. Science 381, 1006-1010.
Kawamoto, N., and Morita, M.T. (2022). Gravity sensing and responses in the coordination of the shoot gravitropic set point angle. New Phytol. 236, 1637-1654.
Kawamoto, N., Kanbe, Y., Nakamura, M., Mori, A., and Morita, M. T. (2020). Gravity-sensing tissues for gravitropism are required for “anti-gravitropic” phenotypes of lzy multiple mutants in Arabidopsis. Plants 9, 615.
Furutani, M., Hirano, Y., Nishimura, T., Nakamura, M., Taniguchi, M., Suzuki, K., Oshida, R., Kondo, C., Sun, S., Kato, K., Fukao, Y., Hakoshima, T., and Morita, M. T. (2020). Polar recruitment of RLD by LAZY1-like protein during gravity signaling in root branch angle control. Nat. Commun. 11, 76.
Taniguchi, M., Furutani, M., Nishimura, T., Nakamura, M., Fushita, T., Iijima, K., Baba, K., Tanaka, H., Toyota, M., Tasaka, M., and Morita, M. T. (2017). Arabidopsis LAZY1 family plays key role in gravity signaling within statocytes in gravitropism and in branch angle control of roots and shoots. Plant Cell 29, 1984-1999.








