基礎生物学研究所



植物が巧みに光を集める仕組みを探る
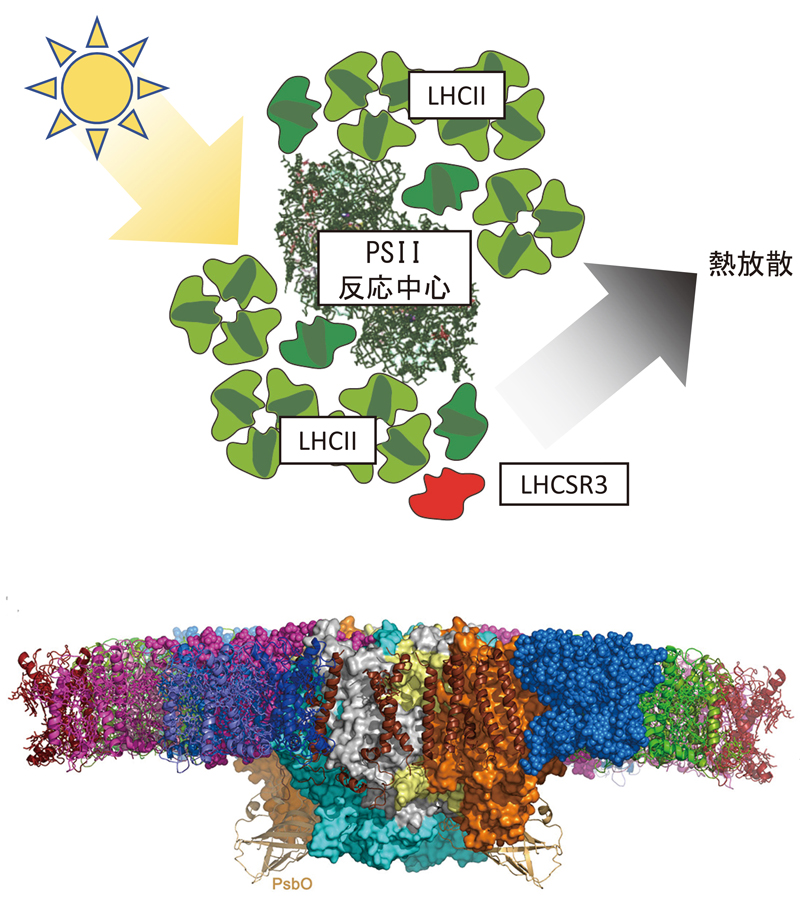
【上】過剰に吸収された光エネルギーを安全に消去するNPQ機構:クラミドモナスの光化学系II(PSII)にLHCSR3が結合すると、集光アンテナ(LHCII)に吸収された光エネルギーはPSII反応中心に移動する前に熱として放散される。このしくみはNPQ(non-photochemical quenching)と呼ばれ、高効率で光を集める光合成装置を強光環境で保護するために役立っている。
【下】原子レベルで解明されたPSII-LHCII超複合体の構造:光化学系IIは、電荷分離を起こす反応中心を光のアンテナであるLHCIIが取り囲んだ構造を取っている。その全体構造がクライオ電顕技術により明らかになった(図はチラコイド膜水平方向からのもの)
植物は、環境の変化に自らを順化適応させることで生き残りをはかる。太陽光を集め、利用可能なエネルギーへの変換を行う光合成においても、さまざまなレベルの光環境適応が行われている。本部門では、単細胞緑藻クラミドモナスを中心としたモデル藻類を用いて、生化学、分子遺伝学、分光学、ライブイメージングなどを駆使し、光合成に必要な光が効率よく集められるしくみや、余分に吸収された光エネルギーを安全に消去するしくみの研究を行っている。
植物や藻類は置かれた環境に応じて光合成装置を変化させ常に最適化された光合成を行っている。その最も顕著な変化は、光を集める“アンテナ”であるLHC(light-harvesting complex)に現れる。本研究部門では、特にLHCに注目し、その光環境適応メカニズムの分子レベルでの解明をめざしている。単細胞緑藻であるクラミドモナスを中心に、さまざまな微細藻類や植物を用い、その光合成装置の先進的な解析を生化学解析、物理学解析、遺伝学解析などを組み合わせて行っている。特に光合成にとって過剰分の光エネルギーを安全に消去する熱放散機構NPQ(non-photochemical quenching)と2つの光化学系へのエネルギー分配機構であるステート遷移に注目し、その分子機構の解明を進めている。私たちは、(1)NPQは、光化学系II超複合体に結合したLHCSRタンパク質が重要であること、(2)LHCSRタンパク質の発現が青色光受容体や紫外線受容体に起因する細胞内シグナル伝達によって起きること、(3)ステート遷移においてリン酸化されたLHCII三量体が光化学系Iに結合する詳細などを明らかにしてきた。最近はクライオ電子顕微鏡を利用した光化学系超複合体の構造解析を足がかりとして、原子レベルで、あるいは膜レベルで光合成装置の環境構造変化を追究している。

図1. 光化学系I超複合体のステート2状態の立体構造
緑藻クラミドモナスを光化学系IIがより励起される状態(ステート2)にしてPSI 超複合体を精製し、クライオ電子顕微鏡画像取得およびコンピュータによる単粒子解析により立体構造を解明した(解像度2.84Å)。超複合体は光化学系Iの片側(図の左側)に光化学系I固有の集光装置であるLhcaの4量体が二層結合し、反対側にLhcaの2量体(Lhca2/9)と光化学系IIの集光装置である3量体LHCII(LHCII-1/LHCII-2)が結合しており多くの微細構造が明らかとなった。特に、ステート2では、LhcbM1とLhcbM5のN末端Thr残基がリン酸化されることで3量体LHCIIがPSIに結合する構造であることが確定した。
本研究部門では、微細藻類を用いて光合成に必要な太陽エネルギーを集める仕組みの研究を行っています。光合成、環境、微細藻類に関心のある方、生命の巧妙な仕組みの解明に挑戦したい方、是非ご連絡ください。基礎知識は問いません。
皆川 純 教授 E-mail: minagawa@nibb.ac.jp








