


2012.07.19 部門公開セミナー
デジタルなDNAメチル化と進化
小林 一三 (東京大学大学院・新領域創成科学研究科・教授)
2012年07月19日(木) 13:30 より 14:30 まで
明大寺地区1階会議室 (111)
生物進化研究部門 長谷部 光泰(7546)
「適応進化は多様な『ゲノム配列』からの選択によって進む」というのが、現在の生命科学の基本的考え方であるが、私は「適応進化は多様な『エピゲノム状態』からの選択によって進む」という考え方を提起したい。根拠は、細菌のDNAメチル化の解析である。
細菌のDNAメチル化酵素の多くは、特異的なDNA配列をメチル化する。それによるゲノムの多数の箇所でのメチル化は、それぞれの近くの遺伝子発現に影響し、グローバルな遺伝子発現のパターンを規定している。DNAメチル化酵素の多くは、制限酵素と共に制限修飾系を作っており、特定の配列でのメチル化というエピジェネティックなIDを持たないDNAは、外からのものであれ、中のものであれ、制限酵素によって破壊される。
細菌ゲノム系列間の遺伝子の動きを妨げる制限修飾系は、それ自身が動く存在である。私たちは、種内ゲノム配列比較から、遺伝子内の「DNA配列認識ドメイン」配列が動く場合を発見した。ある型では、遺伝子内の2つのサイト間で、「DNA配列認識ドメイン」配列が移動していた(図)。「同じ座の遺伝子内で配列が移動する」という、この新しい遺伝子(タンパク質)構造変換のしくみを、「ドメイン移動 (DoMo)」と名付けた。
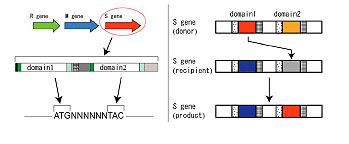
これらから、「一つの細菌が持つ多数のDNAメチル化酵素のそれぞれが、頻繁に認識DNA配列を変換する。それによって、ゲノムのDNAのメチル化状態(メチローム)、グローバルな遺伝子発現、形質が変換する。もたらされる多様なエピゲノムからの選択によって、適応進化が進む」という「エピゲノム駆動進化」仮説を提唱する。
※セミナーは英語で行われます。

