

カツラ科は日本と中国に1属2種が分布。
Two species in one genus in a family are distributed in Japan and China.

前葉は、イネ科など、側枝第一葉の形態が、その後に形成される葉と形態が異なることから名付けられた(Turpin 1819)。双子葉類と裸子植物では側枝の第1葉と第2葉(上図AからC)、単子葉類では側枝の第1葉(上図D)を指す場合が多いが、その後に作られる葉(上図の葉1や葉2など)と形態が異ならない場合が多い(熊澤1979)。双子葉類の場合は2枚の前葉は対生に蓋葉と主軸を結んだ線に直交するように側生し(上図AからC)、単子葉類では向軸側に1枚が生じる場合が多い(上図D)(熊澤 1979)。これら以外にも、いろいろな変異があり、多くの場合、融合や退化によって説明される(熊澤 1979)。双子葉類の場合、2枚の前葉が出た後、互生葉序になる場合(上図A、B)、背軸側(A)と向軸側(B)のそれぞれから次の葉が生じる場合がある。
Turpin, P.I.F. (1819). Mémoire sur l’inflorescence des Graminées et des Cyperées. Mem. Mus. d’Hist. Nat. Paris 5, 426–492.
熊澤正夫(1979)植物器官学、裳華房
 カツラの前葉は右図のように、太い葉脈が2本通っていることから、他の双子葉類に良く見られる側生する2枚の前葉が向軸側で癒合したものではないかと考えられている(熊澤1979)。一方、前葉は腋芽の蓋葉の托葉が癒合してできた可能性もある。あるいは、前葉は向軸側に1枚しかできず、その2枚の托葉が癒合したものである可能性もある(邑田仁博士私信)。カツラの前葉は2枚の前葉が癒合してできているとすると、前葉の葉腋には2個の腋芽が形成される可能性が高い。一方、托葉だとすると腋芽は形成されない。ただ、腋芽が退化消失している可能性もある。従って、2個の腋芽があれば、2枚の前葉が癒合した可能性が高くなるが、腋芽が無い場合は、2枚の前葉が癒合して腋芽が退化した可能性と、托葉が変形したという可能性の両方の可能性が残る。
カツラの前葉は右図のように、太い葉脈が2本通っていることから、他の双子葉類に良く見られる側生する2枚の前葉が向軸側で癒合したものではないかと考えられている(熊澤1979)。一方、前葉は腋芽の蓋葉の托葉が癒合してできた可能性もある。あるいは、前葉は向軸側に1枚しかできず、その2枚の托葉が癒合したものである可能性もある(邑田仁博士私信)。カツラの前葉は2枚の前葉が癒合してできているとすると、前葉の葉腋には2個の腋芽が形成される可能性が高い。一方、托葉だとすると腋芽は形成されない。ただ、腋芽が退化消失している可能性もある。従って、2個の腋芽があれば、2枚の前葉が癒合した可能性が高くなるが、腋芽が無い場合は、2枚の前葉が癒合して腋芽が退化した可能性と、托葉が変形したという可能性の両方の可能性が残る。
Prophylls are the first leaves formed in the axillary shoot. A pair of opposite leaves are usually formed in dicots, while a single leaf in monocots. Morphological diversity of prophylls are explained by fusion or reduction. A prophyll of Cercidiphyllum japonicum has two main veins, suggesting the single prophyll is formed by a fusion of two prophylls (Kumazawa 1979). Another possibility is the fusion of two stipules of a subtending leaf (Dr. Jin Murata, personal communication) .
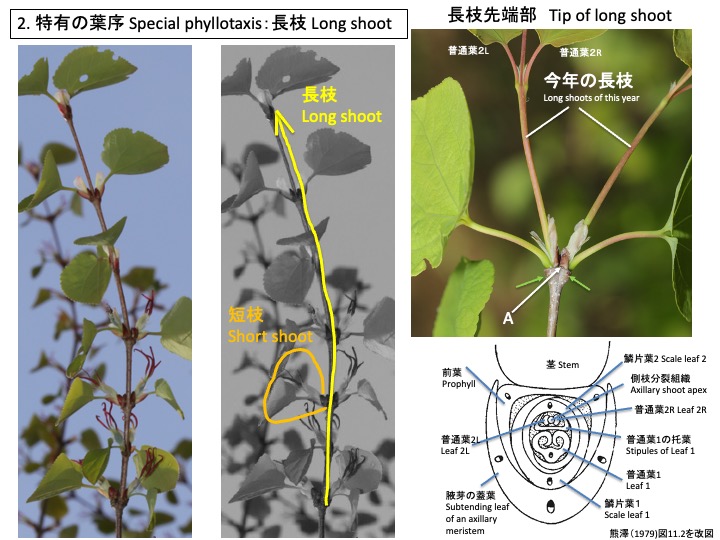 カツラは、長枝と短枝を形成する。ともに仮軸分岐で、前年の葉の腋芽が伸長して新しい年の枝ができる。長枝の場合は、前年の茎頂が枯れ(図のA)、前年に一番先端で対生していた2枚の葉(図の黄緑矢印で落葉した後を示す)のそれぞれの腋芽が伸長する(図に「今年の長枝」と示す)。長枝先端の葉(蓋葉)の葉腋にできた側枝(腋芽)分裂組織は、前葉1枚、鱗片葉2枚、普通葉1枚を茎と蓋葉に直交するように2列互生する(注)。側枝分裂組織は、最初の普通葉(図の普通葉1)の托葉に包まれており、対生する普通葉を次々に形成する。図には、最初の1組の対生普通葉(普通葉2)を示す。最初は2列互生、後に対生に葉序を変化させる点で特有である(熊澤 1979)。
カツラは、長枝と短枝を形成する。ともに仮軸分岐で、前年の葉の腋芽が伸長して新しい年の枝ができる。長枝の場合は、前年の茎頂が枯れ(図のA)、前年に一番先端で対生していた2枚の葉(図の黄緑矢印で落葉した後を示す)のそれぞれの腋芽が伸長する(図に「今年の長枝」と示す)。長枝先端の葉(蓋葉)の葉腋にできた側枝(腋芽)分裂組織は、前葉1枚、鱗片葉2枚、普通葉1枚を茎と蓋葉に直交するように2列互生する(注)。側枝分裂組織は、最初の普通葉(図の普通葉1)の托葉に包まれており、対生する普通葉を次々に形成する。図には、最初の1組の対生普通葉(普通葉2)を示す。最初は2列互生、後に対生に葉序を変化させる点で特有である(熊澤 1979)。
(注)長枝先端の腋芽を観察した報告が見つからなかったので、短枝の断面図(引用した熊澤1979図11.2)、長枝の先端を切ると先端よりの短枝が長枝に変わりうること(Titman and Wetmore 1955) から推定した。
Both long and short shoots of Cercidiphyllum japonicum are sympodial and a shoot of a next year is formed at the leaf axils of the previous year. A prophyll, two scale leaves, and a normal leaf are alternately formed in two rows in both types of shoots. In a long shoot, a lateral shoot meristem covered with stipules of the first normal leaf oppositely form subsequent normal leaves. The switch of alternate phyllotaxis to opposite one is special in Cercidiphyllum and some other angiosperms (Kumazawa 1979).
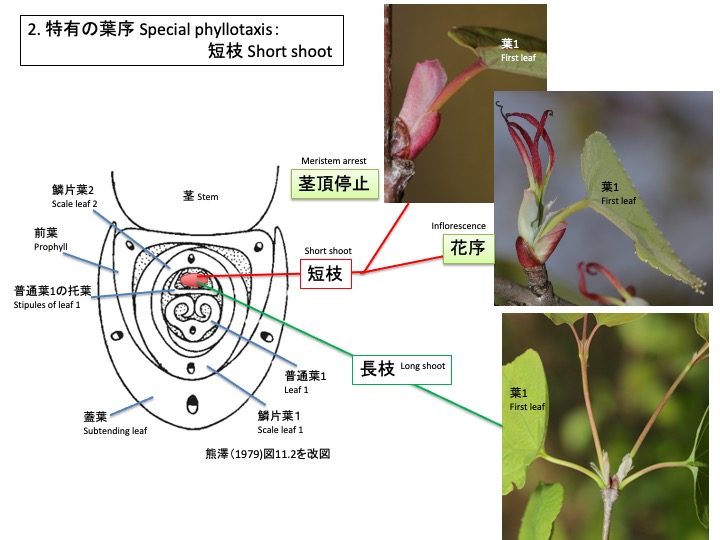 短枝も長枝と同様、蓋葉の葉腋にできた腋芽分裂組織は、前葉1枚、鱗片葉2枚、葉1枚を茎と蓋葉に直交するように2列互生する。長枝との違いは、普通葉1の托葉に包まれた側枝分裂組織が伸長せず、枯れてしまうか、花序となることである。従って、花序をつけない短枝には普通葉は1枚だけ形成される。他の樹木のように短枝に複数の葉を形成せず、1枚だけの葉を形成するため、数年を経た長枝でも、最初に長枝が成長した時のように主軸の両側に葉を対生して見える独特の枝ぶりとなる。
短枝も長枝と同様、蓋葉の葉腋にできた腋芽分裂組織は、前葉1枚、鱗片葉2枚、葉1枚を茎と蓋葉に直交するように2列互生する。長枝との違いは、普通葉1の托葉に包まれた側枝分裂組織が伸長せず、枯れてしまうか、花序となることである。従って、花序をつけない短枝には普通葉は1枚だけ形成される。他の樹木のように短枝に複数の葉を形成せず、1枚だけの葉を形成するため、数年を経た長枝でも、最初に長枝が成長した時のように主軸の両側に葉を対生して見える独特の枝ぶりとなる。
長枝の先端を切除すると直下の短枝は、短枝ではなく長枝を形成する。先端部を切除してもオーキシンを含んだラノリンペーストを塗布すると短枝の長枝化は阻害される(Titman and Wetmore 1955)。このことから、長枝の先端からのオーキシン供給によって下部の腋芽が短枝になるのではないかと推定されている(Titman and Wetmore 1955)。イチョウの短枝と長枝の区別にもオーキシンが関与していると推定されている(Gunckel and Wetmore 1946a, b, Gunckel and Thiman 1949, Gunckel et al. 1949)。シロイヌナズナではオーキシンに加えて、ストリゴラクトンが腋芽の休眠制御に関わっており、カツラやイチョウの場合も関与しているかもしれない。また、カツラは長枝と短枝で葉の枚数が異なっているが、イチョウの短枝は長枝とほぼ同じ数の葉を展開する。展開する葉の枚数を制御する仕組みはわかっていない。
The lateral shoot meristem of a short shoot has two fates. One is to be arrested and the other to form the inflorescence. However, the long shoot meristem is destroyed, lateral shoot meristems of short shoots change their fate to form a long shoot. The difference between long and short shoots should be regulated by auxin (Titman and Wetmore 1955). Similar auxin involvement between long and short shoot development is found in Ginkgo biloba (Gunckel and Wetmore 1946a, b, Gunckel and Thiman 1949, Gunckel et al. 1949). The same number of leaves are formed in a short shoot as that in a long shoot in Ginkgo biloba. Only one leaf is formed in Cercidiphyllum japonicum.
Gunckel, J.E. and Wetmore, R.H. (1946a) Studies of development in long shoots and short shoots of Ginkgo biloba L. I. The origin and pattern of development of the cortex, pith, and procambium. Amer. J. Bot. 33: 285-295.
Gunckel, J.E. and Wetmore, R.H. (1946b) Studies of development in long shoots and short shoots of Ginkgo biloba L. II. Phyl- lotaxis and the organization of the primary vascular system; primary phloem and primary xylem. Amer. J. Bot. 33: 532-543.
Gunckel, J.E. and Thimann, K.V. (1949) Studies of develop- ment in long shoots and short shoots of Ginkgo biloba L. III. Auxin production in shoot growth. Amer. J. Bot. 36: 145-151.
Gunckel, J.E., Thimann, K.V. and Wetmore, R.H. (1949) Studies of development in long shoots and short shoots of Ginkgo biloba L. IV. Growth habit, shoot expression and the mechanism of its control. Amer. J. Bot. 36: 309- 318.
Kumazawa, M. (1979) Plant organography (in Japanese). Shokabo
Titman, P.W. and Wetmore, R.H. (1955) The growth of long and short shoots in Cercidiphyllum. Amer. J. Bot. 42: 354-372.

