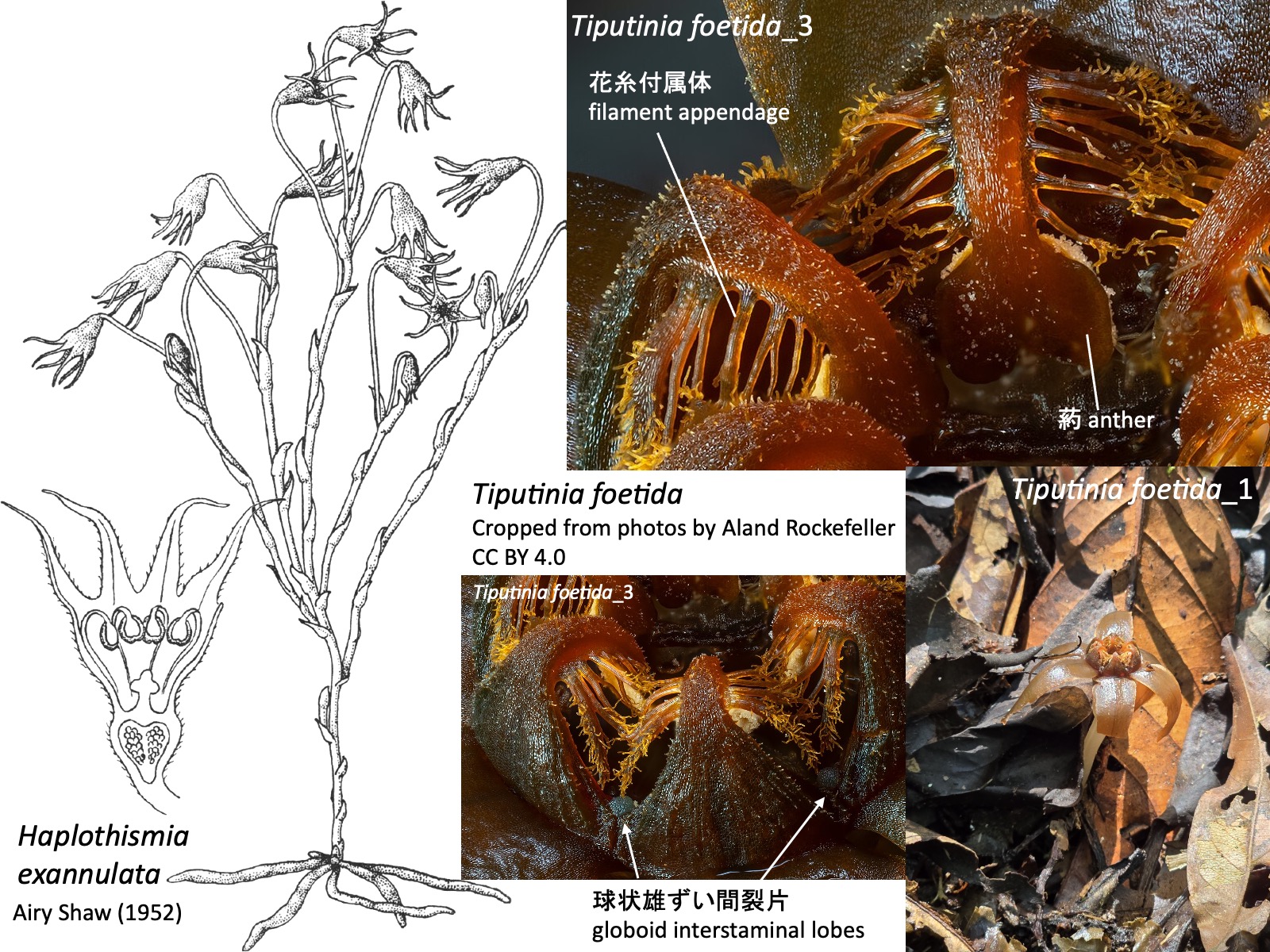
カナボウノキ科 Didiereaceae は、乾燥地に適応した多肉性木本植物からなる。写真は、マダガスカル南部の有刺低木林を代表するAlluaudia proceraである。
The family Didiereaceae consists of succulent woody plants adapted to arid environments. The photograph shows Alluaudia procera, a representative species of the spiny bush forests of southern Madagascar.

カナボウノキ Didierea madagascariensis では、肉質葉が落ちると鋭く強靭な刺だけが残る。倒れると段ボールや絨毯を貫通することもあるため、取り扱いには注意が必要である。
In Didierea madagascariensis, when the succulent leaves fall, only the sharp, robust spines remain. If the plant falls over, the spines may even pierce cardboard or carpets, so it must be handled with care.

Source: POWO (2026). Plants of the World Online. Facilitated by the Royal Botanic Gardens, Kew. Published on the Internet; https://powo.science.kew.org/. Retrieved 7 July 2026. Distribution data from Kew Backbone Distributions (WCVP). © World Checklist of Vascular Plants, licensed under CC BY 3.0.
カナボウノキ科各属の種数と分布 Species number and distribution of genera in Didiereaceae
カナボウノキ科 Didiereaceae は、ポーチュラカリア属 Portulacaria (7種)、カリプトロテカ属 Calyptrotheca (2種)、アラウディオプシス属 Alluaudiopsis (2種)、デカリア属 Decarya (1種)、カナボウノキ属 Didierea (2種)、アラウディア属 Alluaudia (6種)の6属が知られている(POWO)。
The family Didiereaceae comprises six known genera: Portulacaria (7 species), Calyptrotheca (2 species), Alluaudiopsis (2 species), Decarya (1 species), Didierea (2 species), and Alluaudia (6 species) (POWO).

カナボウノキ科の属の系統関係 Phylogeny of Didiereaceae genera
カナボウノキ科 Didiereaceae は、分子系統解析に基づき、アンゴラから南アフリカに分布するポーチュラカリア亜科 Portulacarioideae、東アフリカに分布するカリプトロテカ亜科 Calyptrothecoideae、およびマダガスカルに分布するカナボウノキ亜科 Didiereoideae の3亜科に分かれると推定されている(Bruyns et al. 2014)。
カナボウノキ科は、古第三紀暁新世の約5,900万年前に姉妹群から分岐したと推定されている (Zuntini et al. 2024)。また、ポーチュラカリア属 Portulacaria、カリプトロテカ属 Calyptrotheca がアフリカ大陸に分布すること、さらにマダガスカルが約1億2千万年前にはすでにアフリカ大陸沖の現在の位置にあったことから、カナボウノキ亜科 Didiereoideae は、アフリカ大陸に分布していた祖先が長距離分散によってマダガスカルへ到達し、そこで多様化した可能性が高い (Bruyns 2014)。
Based on molecular phylogenetic analyses, the family Didiereaceae is inferred to comprise three subfamilies: Portulacarioideae, distributed from Angola to South Africa; Calyptrothecoideae, distributed in East Africa; and Didiereoideae, distributed in Madagascar (Bruyns et al. 2014).
The family Didiereaceae is estimated to have diverged from its sister group approximately 59 million years ago, in the Paleocene of the Paleogene (Zuntini et al. 2024). In addition, because Portulacaria and Calyptrotheca occur on the African continent, and because Madagascar had already reached its present position off the coast of Africa by approximately 120 million years ago, the subfamily Didiereoideae most likely originated from an African ancestor that reached Madagascar by long-distance dispersal and subsequently diversified there (Bruyns et al. 2014).

ポーチュラカリア亜科 Portulacarioideae
ポーチュラカリア亜科 Portulacarioideae はポーチュラカリア属 Portulacaria のみを含む。
Portulacarioideae contains only the genus Portulacaria.

Portulacaria afra_1: cropped a photo by Adriaan Grobler, https://www.inaturalist.org/photos/346660774, CC BY-NC
ポーチュラカリア属 – 1. 草姿 Portulacaria – 1: Habit
ポーチュラカリア属 Portulacaria は、アンゴラから南アフリカにかけて自生する7種からなる(POWO)。灌木または低木で、大きくなると高さ数メートルに達する(Carolin 1993)。
The genus Portulacaria comprises seven species native from Angola to South Africa (POWO). They are bushy or shrubby plants, reaching several meters in height when large (Carolin 1993).

ポーチュラカリア属 – 2. 茎葉 Portulacaria – 2. Stems and leaves
茎と葉は肉質である。
The stems and leaves are succulent.

Portulacaria namaquensis_1: cropped from a photo by Karel du Toit (+2782 4928291) , https://www.inaturalist.org/photos/398950896, CC BY-NC
Portulacaria fruticulose_1: cropped from a photo by Karel du Toit (+2782 4928291) , https://www.inaturalist.org/photos/644204588, CC BY-NC
Marloth, R. (1913). The Flora of South Africa, Volume I (Darter Bros. & Co.).

ポーチュラカリア属 – 3. 花序と花 Portulacaria – 3. Inflorescences and Flowers
ガク片は対生する2枚からなり、5枚の花弁は1輪をなして形成される。雄ずいは通常5本で、花弁と互生する(Erbar and Leins 2006)。花は両性花または単性花である(Carolin 1993)。最外側の2枚の花器官が苞葉であるかガク片であるかについては議論がある(Carolin 1993)。カナボウノキ亜科 Didiereoideae では花被が2輪からなるため、最外側の2枚の花器官は苞葉と解釈されている(e.g. Kubitzki 1993)。一方、Portulacaria では花被が1輪であるため、最外側の2枚の花器官がガク片と苞葉のどちらに相同であるかを判断することは難しい。しかし、ここでは Didiereaceae の他の属における苞葉と形態が類似していることから、苞葉として扱う。
単性花を形成する種は、かつて Ceraria 属として区別されていたが(Pearson and Stephens 1912)、分子系統解析により Portulacaria に含まれることが示された(Bruyns et al. 2014)。
The sepals consist of two opposite organs, and the five petals are formed in a single whorl. The stamens are usually five in number and alternate with the petals (Erbar and Leins 2006). The flowers are bisexual or unisexual (Carolin 1993). Whether the two outermost floral organs are bracts or sepals has been debated (Carolin 1993). In the subfamily Didiereoideae, the perianth consists of two whorls, and therefore the two outermost floral organs are interpreted as bracts (e.g. Kubitzki 1993). In Portulacaria, by contrast, the perianth consists of a single whorl, making it difficult to determine whether the two outermost floral organs are homologous to sepals or bracts. Here, however, they are treated as bracts because they are morphologically similar to the bracts in other genera of Didiereaceae.
Species with unisexual flowers were formerly segregated as the genus Ceraria (Pearson and Stephens 1912), but molecular phylogenetic analyses have shown that they are nested within Portulacaria (Bruyns et al. 2014).

Portulacaria afra_3: cropped from a photo by Abdul-lateef Ismail, https://www.inaturalist.org/photos/246678573, CC BY-NC
Portulacaria afra_4: cropped from a photo by Sharon Louw, https://www.inaturalist.org/photos/352168141, CC BY-NC
ポーチュラカリア属 – 4. 果実 Portulacaria – 4. Fruits
果実は薄壁の堅果である。
The fruit is a thin-walled nut.

カリプトロテカ亜科 Calyptrothecoideae
カリプトロテカ亜科 Calyptrothecoideae は、カリプトロテカ属のみを含み、多数の雄ずいをもつこと、および蓋状に開く果実をつけることを特徴とする。
The genus Calyptrotheca is characterized by having numerous stamens and fruits that open by a lid-like cap.

Calyptrotheca taitensis-1: cropped a photo by nsheldrick, https://www.inaturalist.org/photos/616932934, CC BY-NC
Calyptrotheca taitensis-2: cropped a photo by nsheldrick, https://www.inaturalist.org/photos/616932923, CC BY-NC
Calyptrotheca taitensis-3: cropped a photo by nsheldrick, https://www.inaturalist.org/photos/616932905, CC BY-NC
カリプトロテカ属 -1. 草姿と葉 Calyptrotheca -1. Habit and Leaves
カリプトロテカ属 Calyptrotheca は、アフリカ東部に分布する2種からなる (POWO)。低木または小高木で、高さ7 mに達する。葉はやや多肉質で、縁はやや波打つ (Carolin 1993 )。
The genus Calyptrotheca comprises two species distributed in East Africa (POWO). They are shrubs or small trees reaching up to 7 m in height. The leaves are somewhat succulent, with slightly undulate margins (Carolin 1993).

Modified from Pax, F. and Hoffmann, K. (1936), Die natürlichen Pflanzenfamilien, 2nd ed., Bd. 17b: 207. Public domain, via Biodiversity Heritage Library.
Calyptrotheca taitensis-3: cropped a photo by nsheldrick, https://www.inaturalist.org/photos/616932905, CC BY-NC
カリプトロテカ属 -2. 花序と花 Calyptrotheca – 2. Inflorescences and Flowers
カリプトロテカ属 Calyptrotheca の花は、2枚のガク片と5枚の花弁からなる。ただし、最外側の2枚の花器官を苞葉と解釈するか、ガク片と解釈するかについては、Portulacaria の場合と同様に、明確ではない。雄ずいは60本ほどにもなり、1本の花柱が長く伸び出すため、フウチョウソウ科 Capparaceae の花に似ている。この属は、「蓋のある容器」を意味する属名が示すように、蒴果が基部で横裂して6裂し、帽子状の上部が脱落することによって特徴づけられる(上図矢印)。
The flowers of the genus Calyptrotheca consist of two sepals and five petals. However, as discussed for Portulacaria, it is unclear whether the two outermost floral organs should be interpreted as bracts or as sepals. The stamens may number as many as about 60, and a single style elongates prominently, giving the flowers a resemblance to those of Capparaceae. As indicated by the generic name, which means “a lidded container,” this genus is characterized by capsules that split transversely near the base, divide into six lobes, and shed the cap-like upper portion (arrow in the figure above).

Rauh, W. (1983). The morphology and systematic position of the Didiereaceae of Madagascar. Bothalia 14, 839–843. https://doi.org/10.4102/ABC.V14I3/4.1251.
カナボウノキ亜科 Didiereoideae
カナボウノキ亜科 Didiereoideae は、マダガスカル南部から南西部に分布し、トウダイグサ属植物とともに、Euphorbia–Didiereaceae bush と呼ばれる特徴的な有刺低木林(上図の斜線部)を形成する(Rauh 1983)。
The subfamily Didiereoideae is distributed from southern to southwestern Madagascar and, together with species of Euphorbia, forms the characteristic spiny thicket known as Euphorbia–Didiereaceae bush (hatched area in the figure above) (Rauh 1983).

カナボウノキ亜科 Didiereoideae の特徴 Characteristics of the subfamily Didiereoideae
カナボウノキ亜科 Didiereoideae の4属は、長枝の葉腋に生じる腋芽に由来するシュートの発生初期に形成される葉、すなわち初期葉が刺化することを特徴とする。また、4数性の花被および雌雄異株も共有派生形質と考えられる。
刺化する初期葉の位置および数は属によって異なる (Rauh 1956)。アラウディオプシス属 Alluaudiopsis では、短枝となる腋芽において、背軸側の1枚の初期葉が刺化する。デカリア属 Decarya では、腋芽の対生する2枚の初期葉が刺化する。カナボウノキ属 Didierea では、短枝となる腋芽において、数枚の初期葉が刺化する。アルアウディア属 Alluaudia では、短枝となる腋芽において、向軸側の1枚の初期葉が刺化する。
The four genera of the subfamily Didiereoideae are characterized by the modification into spines of leaves formed early in the development of shoots derived from axillary buds in the axils of long-shoot leaves, that is, the early leaves. A tetramerous perianth and dioecy are also considered to be synapomorphies.
The position and number of early leaves modified into spines differ among genera (Rauh 1956). In Alluaudiopsis, a single early leaf on the abaxial side of the axillary bud that develops into a short shoot is modified into a spine. In Decarya, two opposite early leaves of the axillary bud are modified into spines. In Didierea, several early leaves of the axillary bud that develops into a short shoot are modified into spines. In Alluaudia, a single early leaf on the adaxial side of the axillary bud that develops into a short shoot is modified into a spine.

カナボウノキ亜科 Didiereoideae の刺の適応的意義 Adaptive significance of spines in the subfamily Didiereoideae
カナボウノキ亜科 Didiereoideae の刺は、動物による被食に対する防御と考えられる。しかし、写真に示すように、現生のキツネザル類に対しては、必ずしも高い防御効果を示すわけではない。ベレンティ保護区 Berenty Private Reserve では、Lepilemur leucopus が乾季の食物をアルアウディア属 Alluaudia の種の葉に大きく依存している (Crowley and Godfrey 2013)。
カナボウノキ亜科が他の亜科から分岐したのは約1500万年前であり、その後、アフリカからマダガスカルへ祖先が長距離分散し、島内で多様化したと推定されている (Ocampo and Columbus 2010)。その時代のマダガスカルに存在した大型植食動物としては、化石記録に基づき、エピオルニス類 (ゾウチョウ類) とキツネザル類のみが想定されている。エピオルニス類は大きく硬いくちばしをもっていたため、刺による防御効果は限定的であったと考えられる (Crowley and Godfrey 2013)。
そのため、カナボウノキ亜科の初期進化段階では、刺は当時のキツネザル類に対する防御として機能していたが、その後、キツネザル類が刺のある植物を利用できるように進化した可能性があると推定されている (Crowley and Godfrey 2013)。
The spines of the subfamily Didiereoideae are thought to function as a defense against herbivory. However, as shown in the photograph, they do not necessarily provide strong protection against extant lemurs. In Berenty Private Reserve, Lepilemur leucopus relies heavily on the leaves of species of Alluaudia as a food source during the dry season (Crowley and Godfrey 2013).
Didiereoideae is estimated to have diverged from the other subfamilies approximately 15 million years ago, after which its ancestor dispersed from Africa to Madagascar by long-distance dispersal and diversified on the island (Ocampo and Columbus 2010). Based on the fossil record, the only large herbivores inferred to have been present in Madagascar at that time were elephant birds and lemurs. Because elephant birds had large, hard beaks, the defensive effect of spines against them is thought to have been limited (Crowley and Godfrey 2013). Thus, it has been suggested that, during the early evolution of Didiereoideae, the spines functioned as a defense against lemurs present at that time, but that lemurs subsequently evolved the ability to exploit spiny plants (Crowley and Godfrey 2013).

Alluaudiopsis fiherenensis_1: cropped a photo by Solofo Eric Rakotoarisoa, https://www.inaturalist.org/photos/106738053, CC BY-NC
Alluaudia comosa_1: cropped a photo by Joshua Stone, https://www.inaturalist.org/photos/675711888, CC BY-NC
Decarya madagascariensis_1: cropped a photo by Franck Rakotonasolo, https://www.inaturalist.org/photos/2041841, CC BY-NC
カナボウノキ亜科 Didiereoideae の藪状形態の適応的意義 Adaptive significance of the bushy habit in the subfamily Didiereoideae
アラウディオプシス属 Alluaudiopsis とアルアウディア属 Alluaudia の種は、生育初期には弾力性のある枝をもち、広角に分枝する藪状の形態を示す。また、デカリア属 Decarya の種は、成長後も藪状の形態を保つ。これらの形態は、陸ガメ類、カバ類、キツネザル類などの植食動物に対しては、それほど高い防御効果を示さないと考えられる (Crowley and Godfrey 2013)。一方、絶滅したエピオルニス類にとっては、大きなくちばしで弾力性のある枝の藪の中から葉を食べることは困難であり、防御効果があったと考えられている(Bond and Silander 2007)。現在では、エピオルニス類の絶滅に伴い、これらの形態は進化的アナクロニズム、すなわち絶滅した生物との相互作用に由来する適応形質の名残である可能性があると考えられている (Crowley and Godfrey 2011; Bond and Silander 2007)。同様に、大型の飛べない鳥類であるモア類が絶滅したニュージーランドでも、似た枝ぶりをもつ植物が見られ、それらも進化的アナクロニズムと考えられている (Bond and Silander 2007)。
Species of Alluaudiopsis and Alluaudia have elastic branches during early growth and show a bushy habit with widely divergent branching. Species of Decarya also retain a bushy habit even when mature. These forms are thought to provide relatively little protection against herbivores such as tortoises, hippos, and lemurs (Crowley and Godfrey 2013). By contrast, for the extinct elephant birds, it would have been difficult to feed on leaves from within a thicket of elastic branches using their large beaks, and these forms are therefore thought to have had a defensive function against them (Bond and Silander 2007). Today, following the extinction of elephant birds, these forms may represent evolutionary anachronisms, that is, remnants of adaptive traits derived from interactions with extinct organisms (Crowley and Godfrey 2011; Bond and Silander 2007). Similarly, in New Zealand, where moas, large flightless birds, have gone extinct, plants with similar branching patterns are found and are also regarded as evolutionary anachronisms (Bond and Silander 2007).

アルアウディオプシス属 Alluaudiopsis の特徴 Characteristics of the genus Alluaudiopsis
アルアウディオプシス属 Alluaudiopsis は、長枝の葉腋に生じる腋芽において、背軸側、すなわち長枝の基部側に位置する1枚の初期葉が刺化することを特徴とする。
The genus Alluaudiopsis is characterized by the modification into a spine of a single early leaf located on the abaxial side of the axillary bud formed in the axil of a long-shoot leaf.

Alluaudiopsis fiherenensis_1: cropped a photo by CORDENOS Thierry, https://www.inaturalist.org/photos/463788613, CC BY-NC
Alluaudiopsis marnieriana_1: cropped a photo by Rosario Douglas, https://www.inaturalist.org/photos/438170892, CC BY
アルアウディオプシス属 Alluaudiopsis – 1. 草姿 Alluaudiopsis – 1. Habit
アルアウディオプシス属 Alluaudiopsis は、マダガスカル南部から南西部に分布する2種からなる(POWO)。藪状または低木状の植物である (Kubitzki 1993)。
The genus Alluaudiopsis comprises two species distributed from southern to southwestern Madagascar (POWO). They are bushy or shrubby plants (Kubitzki 1993).

アルアウディオプシス属 Alluaudiopsis – 2. 茎葉 Alluaudiopsis – 2. Stems and Leaves
アルアウディオプシス属 Alluaudiopsis の種では、長枝につく肉質葉(白色矢印)の腋芽が短枝となる。その短枝では、背軸側、すなわち長枝の基部側に位置する1枚の初期葉が刺(青色矢印)へと変形するため、短枝の頂端分裂組織(赤色矢印)は刺よりも長枝の先端側に位置する。この頂端分裂組織から、肉質の栄養葉(橙色矢印)が形成される。
In species of Alluaudiopsis, the axillary bud of a succulent leaf on the long shoot (white arrows) develops into a short shoot. In this short shoot, a single early leaf located on the abaxial side, that is, toward the base of the long shoot, is modified into a spine (blue arrows). As a result, the apical meristem of the short shoot (red arrows) is positioned distal to the spine, toward the apex of the long shoot. Succulent foliage leaves (orange arrows) are formed from this apical meristem.

長枝の葉が落葉した後は、生育期ごとに短枝から新しい葉が形成される。写真に見える葉はすべて短枝の葉である。
After the leaves on the long shoots fall, new leaves are produced from the short shoots during each growing season. All the leaves visible in the photograph are short-shoot leaves.

Alluaudiopsis marnieriana_2: cropped a photo by Rob C. H. M. Oudejans, https://www.inaturalist.org/photos/58429619, CC BY-NC
Alluaudiopsis marnieriana_3: cropped a photo by Laurence Ramon, https://www.inaturalist.org/photos/399348908, CC BY-NC
Alluaudiopsis marnieriana_4: cropped a photo by Andy Goerdel – Texas Master Naturalist, Good Water Chapter, https://www.inaturalist.org/photos/586700401, CC BY-NC
Alluaudiopsis fiherenensis_3: cropped a photo by nomentsoa, https://www.inaturalist.org/photos/592575626, CC BY-NC
Rauh, W. (1963). 121 Famille. Didiereacees. In Flore de Madagascar et des Comores: plantes vasculaires, , H. Humbert and J.-F. Leroy, eds. (Museum National D’Histoire Naturelle). CC BY-NC-SA 4.0
アルアウディオプシス属 Alluaudiopsis – 3. 花序と花 Alluaudiopsis – 3. Inflorescences and Flowers
カナボウノキ亜科 Didiereoideae の4属はいずれも、右上写真に示すように、対生する苞葉をもち、2枚の外花被片と2枚の内花被片を形成する(Erbar and Leins 2006)。ポーチュラカリア亜科 Portulacarioideae およびカリプトロテカ亜科 Calyptrothecoideae に見られる5数性の花被が祖先形質であり、カナボウノキ亜科の共通祖先において、各輪2数性の花被が進化したと推定されている(Erbar and Leins 2006)。雄ずいは6–12本で1輪をなし、そのうち2本が外花被片と、別の2本が内花被片と対生することが多い(Erbar and Leins 2006)。苞葉は宿存性で、果実を包む。上図の Alluaudiopsis marnieriana の果実図(Rauh 1963)では、苞葉が取り除かれている。
All four genera of the subfamily Didiereoideae have opposite bracts and form two outer and two inner perianth segments, as shown in the upper right of the figure above (Erbar and Leins 2006). The pentamerous perianth found in Portulacarioideae and Calyptrothecoideae is inferred to represent the ancestral condition, and a perianth with two segments in each whorl is thought to have evolved in the common ancestor of Didiereoideae (Erbar and Leins 2006). The stamens number 6–12 and form a single whorl; two of them are often opposite the outer perianth segments and another two are often opposite the inner perianth segments (Erbar and Leins 2006). The bracts are persistent and enclose the fruit. In the fruit illustration of Alluaudiopsis marnieriana shown above (Rauh 1963), the bracts have been removed.

デカリア属 Decarya の特徴 Characteristics of the genus Decarya
デカリア属 Decarya では、長枝の葉腋に生じる腋芽において、対生する2枚の初期葉が刺化する。腋芽は長枝として伸長する場合と、伸長せずに活動を停止する場合があり、他のカナボウノキ亜科の属に見られるような典型的な短枝は形成しない。また、長枝がジグザグ状に屈曲することも、この属を特徴づける形質である。
In the genus Decarya, two opposite early leaves of the axillary bud formed in the axil of a long-shoot leaf are modified into spines. The axillary bud may elongate as a long shoot or cease activity without elongating, and it does not form a typical short shoot like those seen in other genera of Didiereoideae. The zigzag bending of the long shoots is also a characteristic feature of this genus.

Decarya madagascariensis_1: cropped from a photo by mamy_andriamahay, https://www.inaturalist.org/photos/9358854, CC BY-NC
Decarya madagascariensis_2: cropped from a photo by Laurence Ramon, https://www.inaturalist.org/photos/511816461, CC BY-NC
デカリア属 Decarya – 1. 草姿 Decarya – 1. Habit
デカリア属 Decarya は、マダガスカル南部に分布する1種のみからなる (POWO)。藪状、低木状、または小高木状になる (Kubitzki 1993)。
The genus Decarya comprises a single species distributed in southern Madagascar (POWO). It is bushy, shrubby, or small-tree-like in habit (Kubitzki 1993).

デカリア属 Decarya – 2. 茎葉 Decarya – 2. Stems and Leaves
デカリア属 Decarya の種では、長枝につく肉質葉(白色矢印)は早落性であり、その葉腋に腋芽が形成される。この腋芽では、対生する2枚の初期葉が刺 (青色矢印) へと変形するため、腋芽の頂端分裂組織 (赤色矢印) は2本の刺の間に位置する。デカリア属の刺は長枝の葉の托葉のようにも見えるが、カナボウノキ科の他の属では托葉が形成されないこと、刺が長枝の葉の発生後に生じること、また刺が長枝の葉の基部よりも茎頂側に形成されることから、腋芽に由来する初期葉であると考えられている (Rauh 1960/1961)。
デカリア属では、長枝の葉は互生するが、腋芽に由来する刺は対生する。腋芽の初期葉のうち、最初に形成される1枚または数枚は前出葉 prophyll と呼ばれ (Choob 2022)、その後に形成される葉とは葉序や形態が異なることがある。そのため、デカリア属の刺は前出葉である可能性がある。2枚の刺葉を形成した後、腋芽は長枝として伸長するか、または活動を停止し、それ以降の葉を形成しない (Rauh 1960/1961)。
In species of Decarya, the succulent leaves on the long shoots (white arrows) are caducous, and axillary buds are formed in their axils. In these buds, two opposite early leaves are modified into spines (blue arrows), so that the apical meristem of the axillary bud (red arrow) is positioned between the two spines. Although the spines of Decarya may appear similar to stipules of the long-shoot leaves, they are considered to be early leaves derived from axillary buds because stipules are not formed in other genera of Didiereaceae, because the spines arise after the initiation of the long-shoot leaves, and because they are formed distal to the bases of the long-shoot leaves, toward the shoot apex (Rauh 1960/1961).
In Decarya, the leaves on the long shoots are alternate, whereas the spines derived from the axillary buds are opposite. The first one or several early leaves of an axillary bud are called prophylls (Choob 2022), and they may differ in phyllotaxis and morphology from leaves formed later. Therefore, the spines of Decarya may represent prophylls. After forming two spine leaves, the axillary bud either elongates as a long shoot or ceases activity and forms no further leaves (Rauh 1960/1961).

デカリア属 Decarya は、上の写真のように、長枝の形成時に枝がジグザグ状に伸び、成長後もその形状を維持する。
デカリア属のジグザグ状シュートの適応的意義は、まだ実証されていない。先述したように、藪状の形態をつくることで、乾燥地環境における被食防御として機能している可能性がある。また、長枝葉が短命で、一年の多くの期間を無葉で過ごすことを考えると、ジグザグ状の枝は、茎による光合成面積の確保に寄与している可能性もある。ジグザグ状の枝が発生的にどのように形成されるのかについては、まだ調べられていない。
In the genus Decarya, as shown in the photograph above, branches elongate in a zigzag pattern during the formation of long shoots and retain this shape after maturation. The adaptive significance of the zigzag shoots in Decarya has not yet been demonstrated. As discussed above, they may function as a defense against herbivory in arid environments by forming a bushy habit. In addition, because the long-shoot leaves are short-lived and the plants remain leafless for much of the year, the zigzag branches may also contribute to maintaining photosynthetic surface area in the stems. How these zigzag branches are formed developmentally has not yet been investigated.

Decarya madagascariensis_3: cropped from a photo by mamy_andriamahay, https://www.inaturalist.org/photos/9358854, CC BY-NC
Rauh, W. (1963). 121 Famille. Didiereacees. In Flore de Madagascar et des Comores: plantes vasculaires, , H. Humbert and J.-F. Leroy, eds. (Museum National D’Histoire Naturelle). CC BY-NC-SA 4.0
デカリア属 Decarya – 3. 花序と花 Decarya – 3. Inflorescences and Flowers
デカリア属 Decarya の種では、花は分枝した集散花序につく。他のカナボウノキ亜科の種と同様に、対生する2枚の苞葉、2枚の外花被片と2枚の内花被片、8本程度の雄ずい、および1本の雌ずいを形成する(Rauh 1963)。雄花では雌ずいが、雌花では雄ずいが成熟しない(Rauh 1963)。
In species of Decarya, the flowers are borne in branched cymose inflorescences. As in other species of Didiereoideae, each flower forms two opposite bracts, two outer and two inner perianth segments, about eight stamens, and a single pistil (Rauh 1963). In male flowers, the pistil does not mature, whereas in female flowers, the stamens do not mature (Rauh 1963).

カナボウノキ属 Didierea の特徴 Characteristics of the genus Didierea
カナボウノキ属 Didierea では、長枝の葉腋に生じる腋芽において、数枚の初期葉が刺化し、茎の髄が発達し、貯水組織として機能することを特徴とする。
The genus Didierea is characterized by the modification of several early leaves into spines in axillary buds formed in the axils of long-shoot leaves, and by the development of the stem pith as a water-storage tissue.

Didierea madagascariensis_1: cropped a photo by Nick Leggatt, https://www.inaturalist.org/photos/681559740, CC BY-NC
Didierea trollii_1: cropped a photo by Solofo Eric Rakotoarisoa, https://www.inaturalist.org/photos/18818029, CC BY-NC
カナボウノキ属 Didierea – 1. 草姿 Didierea – 1. Habit
カナボウノキ属 Didierea は、マダガスカル南部から南西部に分布する2種からなる (POWO)。高さ数m程度に達する灌木または低木である (Kubitzki 1993)。D. trollii は、若い時期に匍匐性または斜上性の枝(右図矢印)を形成する特徴がある (Rauh 1963)。
The genus Didierea comprises two species distributed from southern to southwestern Madagascar (POWO). They are bushy or shrubby plants reaching several meters in height (Kubitzki 1993). D. trollii is characterized by the formation of creeping or ascending branches during its juvenile stage (arrow in the figure on the right) (Rauh 1963).

カナボウノキ属 Didierea – 2. 茎葉 Didierea – 2. Stems and Leaves
カナボウノキ属 Didierea の種では、長枝につく肉質葉 (白色矢印) の腋芽が短枝となる。この短枝に生じる数枚の初期葉は刺 (青色矢印) へと変形し、その後に肉質葉 (橙色矢印) が形成される。写真では刺は3本見えるが、12本ほど形成されることもある (Kubitzki 1993)。短枝の頂端分裂組織は、これらの刺や葉の中央に位置する。刺は腋芽が伸長する最初の年にだけ形成される。肉質葉は乾季に落葉し、雨季になると短枝の頂端分裂組織から数枚の栄養葉が形成される (Rauh 1983)。
In species of Didierea, the axillary buds of the succulent leaves on the long shoots (white arrow) develop into short shoots. Several early leaves formed on these short shoots are modified into spines (blue arrows), and succulent leaves (orange arrows) are formed subsequently. Although three spines are visible in the photograph, as many as about 12 may be formed (Kubitzki 1993). The apical meristem of the short shoot is located at the center of these spines and leaves. Spines are formed only in the first year of axillary-bud elongation. The succulent leaves fall during the dry season, and when the rainy season begins, several vegetative leaves are formed from the apical meristem of the short shoot (Rauh 1983).

長枝の肉質葉 (黄色矢印) のすべての葉腋から短枝が形成される。長枝の肉質葉は早落性であり、写真ではすでにほとんどが落葉して、葉痕 (白色矢印) が見えている。翌年以降の雨季には、短枝の葉のみが形成される。
Short shoots are formed in all axils of the succulent leaves on the long shoots (yellow arrow). The succulent leaves on the long shoots are caducous, and in the photograph most have already fallen, leaving visible leaf scars (white arrows). In the rainy seasons from the following year onward, only leaves of the short shoots are formed.

短枝を茎頂側から見た様子 (左写真)。短枝の頂端分裂組織は、刺や葉に囲まれた中央部に位置する。前出葉である刺と、その後に形成される肉質の栄養葉は、ともに葉序を構成する。茎頂分裂組織 (右写真、黒色矢印) は、葉基の奥に沈み込んでいる。
Short shoot viewed from the apical side (left photograph). The apical meristem of the short shoot is located in the central region surrounded by the spines and leaves. The spines, which are prophylls, and the succulent vegetative leaves formed subsequently together constitute the phyllotaxis. The shoot apical meristem (right photograph, black arrow) is sunken deep among the leaf bases.

カナボウノキ属 Didierea に特徴的な強靭な刺は、表皮細胞と表皮下細胞が硬化することで形成される (Rauh 1956)。硬化した外層を取り除くと、刺は鞘状の構造をしていることがわかる。その内側には緑色の皮層があり、硬化層を透過した光を利用して光合成を行っている可能性がある。
The robust spines characteristic of the genus Didierea are formed by the hardening of epidermal and subepidermal cells (Rauh 1956). When the hardened outer layer is removed, the spine is seen to have a sheath-like structure. Inside this layer is a green cortex, which may carry out photosynthesis using light transmitted through the hardened layer.

カナボウノキ Didierea madagascariensis の長枝の先端付近を切断すると (左図の赤色点線)、外側から、硬化した表層組織、緑色の皮層、やや木質化した木部を含む維管束組織、そして茎の大部分を占める髄からなることがわかる。茎の内部には樹脂組織 (黒色矢印) があり、切断すると粘性のある透明な液体が滲み出す。
When a long shoot of Didierea madagascariensis is cut near its apex (red dashed line in the left figure), it can be seen to consist, from the outside inward, of a hardened superficial tissue, a green cortex, vascular tissue containing slightly lignified xylem, and a pith that occupies most of the stem. Resinous tissue is present inside the stem (black arrow), and when the stem is cut, a viscous, transparent liquid exudes from it.

茎の先端から約1 m基部側を切断すると (左写真の赤色実線)、髄にははしご状の隙間が認められる。先端付近ではこのような隙間は認められないため、成長過程で形成されると考えられる。隙間の表面は平滑で、茶色の細胞壁をもつ細胞が点在しているが、その形成過程は明らかにされていない。髄は水分貯蔵組織として機能すると考えられており、乾季には髄から皮層や茎頂へ水分が供給されると考えられる。その際、髄に切れ目があることで、水分の減少に伴って組織が収縮しても、組織全体が断裂することを防いでいるのかもしれない。
When the stem is cut about 1 m below the apex (red solid line in the left photograph), ladder-like spaces are observed in the pith. Because such spaces are not observed near the apex, they are thought to form during the course of growth. The surfaces of the spaces are smooth, and cells with brown cell walls are scattered on them, but the process by which these spaces form remains unclear. The pith is thought to function as a water-storage tissue, and during the dry season, water is presumably supplied from the pith to the cortex and the shoot apex. The presence of breaks in the pith may prevent the entire tissue from rupturing as the tissue contracts in response to water loss.

長枝を切断した後、切断面 (左写真、赤色実線) に露出した髄は約2週間で乾燥した (中央写真)。一方、切断後約2か月半が経過してから長枝の先端部を再度切断すると (左写真、赤色点線)、内部はなお水分を保持していた。このことから、茎は高い水分保持能力をもつと考えられる。
After the long shoot was cut, the pith exposed on the cut surface (left photograph, red solid line) dried out in about two weeks (middle photograph). By contrast, when the apical part of the long shoot was cut again about two and a half months after the initial cut (left photograph, red dashed line), the interior still retained water. This suggests that the stem has a high capacity for water retention.

皮層や髄には、透明に見える分泌組織 (黒色矢印) が散在する。分泌物は粘性をもち、傷害時に傷口の保護に関与している可能性がある。
Secretory tissues that appear transparent (black arrows) are scattered in the cortex and pith. The secretion is viscous and may be involved in protecting wounds after injury.

髄に隙間がまだ形成されていない茎の先端部を切断すると (左上写真)、切り口の組織が変化し、褐色で乾燥した硬い膜状の枯死組織が形成される (右上写真。左上写真とは別の切断面を示す)。この枯死組織により、内側の組織は乾燥から保護されていると考えられる (左下写真)。髄に隙間がある部分で切断した場合にも、同様の枯死組織が形成され、内部を保護している (右下写真。枯死組織を取り除き内側を示したもの)。
When the apical part of a stem in which spaces have not yet formed in the pith is cut (upper left photograph), the tissue at the cut surface changes and forms a brown, dry, hard, membrane-like necrotic tissue (upper right photograph, showing a different cut surface from that in the upper left photograph). This necrotic tissue is thought to protect the inner tissues from desiccation (lower left photograph). When the stem is cut in a region where spaces are present in the pith, similar necrotic tissue is also formed and protects the interior (lower right photograph, showing the inside after the necrotic tissue was removed).

Didierea madagascariensis_2: cropped a photo by avocat, https://www.inaturalist.org/photos/297193648, CC BY
Didierea trollii_2: cropped a photo by CORDENOS Thierry, https://www.inaturalist.org/photos/77696178, CC BY-NC
Rauh, W. (1963). 121 Famille. Didiereacees. In Flore de Madagascar et des Comores: plantes vasculaires, , H. Humbert and J.-F. Leroy, eds. (Museum National D’Histoire Naturelle), CC BY-NC-SA 4.0

Didierea trollii_3: cropped a photo by dennis-mada, https://www.inaturalist.org/photos/4727865, CC BY-NC
カナボウノキ属 Didierea – 3. 花序と花 Didierea – 3. Inflorescences and Flowers
カナボウノキ属の花は、短枝の頂端に束生する。
The flowers of the genus Didierea are borne in clusters at the apices of short shoots.

Didierea trollii_4: cropped a photo by dennis-mada, https://www.inaturalist.org/photos/4727865, CC BY-NC
Rauh, W. (1963). 121 Famille. Didiereacees. In Flore de Madagascar et des Comores: plantes vasculaires, , H. Humbert and J.-F. Leroy, eds. (Museum National D’Histoire Naturelle). CC BY-NC-SA 4.0
カナボウノキ属 Didierea の花は、他のカナボウノキ亜科の種と同様に、対生する2枚の苞葉、2枚の外花被片と2枚の内花被片、8本程度の雄ずい、および1本の雌ずいを形成する (Rauh 1963)。雄花では雌ずいが、雌花では雄ずいが形成されるが、いずれも成熟しない (Rauh 1963)。
The flowers of the genus Didierea, as in other species of Didiereoideae, form two opposite bracts, two outer and two inner perianth segments, about eight stamens, and a single pistil (Rauh 1963). In male flowers, the pistil is formed but does not mature, whereas in female flowers, the stamens are formed but do not mature (Rauh 1963).

アルアウディア属 Alluaudia の特徴 Characteristics of the genus Alluaudia
アルアウディア属 Alluaudia は、長枝の葉腋に生じる腋芽において、向軸側、すなわち長枝の先端側に位置する1枚の初期葉が刺化することを特徴とする。また、Alluaudia dumosa 以外の種では、短枝に対生する2枚の肉質葉を形成する。
The genus Alluaudia is characterized by the modification into a spine of a single early leaf located on the adaxial side, that is, toward the apex of the long shoot, in the axillary bud formed in the axil of a long-shoot leaf. In species other than Alluaudia dumosa, two opposite succulent leaves are formed on the short shoots.

Alluaudia comosa_1: cropped a photo by Laurence Ramon, https://www.inaturalist.org/511786677, CC BY-NC
アルアウディア属 Alluaudia – 1. 草姿 Alluaudia – 1. Habit
アルアウディア属 Alluaudia は、マダガスカル南部に自生する6種からなる (POWO, GBIF.org, accessed 13 July 2026)。高さ5 m程度の灌木状または低木状の種から、高さ10 mほどの高木状になる種まで含む (Rauh 1963)。Alluaudia dumosa を除き、短枝に対生する2枚程度の葉をつけることを特徴とする (左上写真)。
The genus Alluaudia comprises six species native to southern Madagascar (POWO, GBIF.org, accessed 13 July 2026). It includes bushy or shrubby species reaching about 5 m in height, as well as tree-like species reaching about 10 m (Rauh 1963). With the exception of Alluaudia dumosa, the genus is characterized by the formation of about two opposite leaves on the short shoots (upper left photograph).


アルアウディア属 Alluaudia – 2. 茎葉 Alluaudia – 2. Stems and Leaves
アルアウディア属 Alluaudia の種では、長枝につく肉質葉(白色矢印)の腋芽が短枝となる。この短枝では、向軸側、すなわち長枝の先端側に位置する1枚の初期葉が刺(青色矢印)へと変形する。長枝の茎頂付近では、短枝の茎頂分裂組織は組織に埋もれて外部からは見えず、長枝の肉質葉の葉腋から直接刺が生じているように見える(上中央写真)。長枝が成長するにつれて、介在分裂により長枝の肉質葉と刺の間の組織が伸長し(黒色両矢印)、両者が離れると短枝の茎頂(赤色矢印)が見えるようになる(右上写真)。さらに長枝が成長すると、短枝の茎頂から対生する2枚の肉質葉(橙色矢印)が形成される。
In species of Alluaudia, the axillary buds of the succulent leaves on the long shoots (white arrow) develop into short shoots. In these short shoots, a single early leaf located on the adaxial side, that is, toward the apex of the long shoot, is modified into a spine (blue arrow). Near the shoot apex of the long shoot, the shoot apical meristem of the short shoot is embedded in the tissue and is not visible externally, making it appear as though the spine arises directly from the axil of the succulent leaf on the long shoot (upper middle photograph). As the long shoot grows, the tissue between the succulent leaf of the long shoot and the spine elongates through intercalary growth (black double-headed arrow), and when the two become separated, the apex of the short shoot (red arrow) becomes visible (upper right photograph). As the long shoot grows further, two opposite succulent leaves (orange arrow) are formed from the apex of the short shoot.

乾季には長枝および短枝の肉質葉が落葉するが、翌年以降に新たに形成されるのは短枝の肉質葉のみである。
During the dry season, the succulent leaves of both long and short shoots fall, but from the following year onward, only the succulent leaves of the short shoots are newly formed.

左の写真は、倒れた Alluaudia procera の茎の横断面である。木部 (茶色矢印) の内側には髄 (水色両矢印) が、外側には皮層 (緑色両矢印) が位置する。双子葉植物における一次肥大成長は、皮層または髄の細胞増殖によって引き起こされるとされており (Troll and Rauh 1950)、アルアウディア属 Alluaudia やサボテン科では皮層が肥大成長する(右上模式図左)のに対し、カナボウノキ属 Didierea では髄が肥大成長する (右上模式図右) (Rauh 1956)。
The photograph on the left shows a transverse section of the stem of a fallen Alluaudia procera. The pith (light blue double-headed arrow) is located inside the xylem (brown arrow), and the cortex (green double-headed arrow) is located outside it. Primary thickening growth in dicotyledons is thought to be caused by cell proliferation in either the cortex or the pith (Troll and Rauh 1950). In the genus Alluaudia and the family Cactaceae, the cortex undergoes thickening growth (upper right schematic, left), whereas in the genus Didierea, the pith undergoes thickening growth (upper right schematic, right) (Rauh 1956).

Alluaudia comosa_2: cropped a photo by globalherping, https://www.inaturalist.org/photos/401384720, CC BY-NC
Alluaudia comosa_3: cropped a photo by Louis Aureglia, https://www.inaturalist.org/photos/455583401, CC BY
アルアウディア属 Alluaudia – 3. 花序と花 Alluaudia – 3. Inflorescences and Flowers
アルアウディア属 Alluaudia の花は、他のカナボウノキ亜科の種と同様に、対生する2枚の苞葉、2枚の外花被片と2枚の内花被片、8本程度の雄ずい、および1本の雌ずいを形成する (Rauh 1963)。雄花では雌ずいが、雌花では雄ずいが形成されるが、いずれも成熟しない (Rauh 1963)。
The flowers of the genus Alluaudia, as in other species of Didiereoideae, form two opposite bracts, two outer and two inner perianth segments, about eight stamens, and a single pistil (Rauh 1963). In male flowers, the pistil is formed but does not mature, whereas in female flowers, the stamens are formed but do not mature (Rauh 1963).
Alluaudia の種への検索表 (Rauh 1963を改変)
アルアウディア属 Alluaudia では、種によって、草丈、枝分かれの程度、短枝における肉質葉の有無、花序の形態、および雌花の苞葉に形成される翼の形態が異なる。
1. 短枝に肉質の葉を形成しない。 …… Alluaudia dumosa
1. 短枝に肉質の葉を形成する。 …… 2
2. 花序軸は発達せず、花は長枝の葉腋に束生する。果時に雌花の苞葉背面に翼が発達する。樹冠の頂部で長枝が活発に分枝し、枝が密集する。 …… Alluaudia comosa
2. 花序軸は発達し、花は長枝の葉腋から出る房状の花序に付く。果時に雌花の苞葉背面に翼が発達しない。樹冠の頂部で長枝はそれほど活発には分枝しない。…… 3
3. 花序は大型で、直径30 cmに達し、枝先近くに集散花序となる。 …… 4
3. 花序はより小型で、長さおよび幅は最大15 cm。長枝の上部に段階的につく。 …… 5
4. 幼植物は基部からよく分枝する。成木の側枝はかなり多い。短枝の葉は倒卵形で、先端は丸く、若い時期には有毛。 …… Alluaudia procera
4. 幼植物は基部から分枝しない。成木の側枝は少ない。短枝の葉は円形で、先端は切れ込む。無毛。 …… Alluaudia montagnacii
5. 高さ12 mに達する高木で、幹は太く、側枝は直立し少数。短枝の葉は円形で、先端は切れ込む。雌花の苞葉の翼の大きさは、両方の苞葉でほぼ同じ。 …… Alluaudia ascendens
5. 高さ5 mに達する低木または小高木で、幹は細く、側枝は多数あり、細長く、多少開出する。短枝の葉は倒卵形で、先端は丸いか、切形またはわずかに凹む。雌花の苞葉の翼の大きさは、片方の苞葉が他方より顕著に大きい。 …… Alluaudia humbertii
Key to the species of Alluaudia (modified from Rauh 1963)
In the genus Alluaudia, species differ in plant height, degree of branching, presence or absence of succulent leaves on short shoots, inflorescence morphology, and the morphology of the wings formed on the bracts of female flowers.
1. Succulent leaves are not formed on the short shoots. …… Alluaudia dumosa
1. Succulent leaves are formed on the short shoots. …… 2
2. The inflorescence axis is not developed, and the flowers are clustered in the axils of long-shoot leaves. In fruit, wings develop on the dorsal surface of the bracts of the female flowers. Long shoots branch actively at the top of the crown, where the branches are densely crowded. …… Alluaudia comosa
2. The inflorescence axis is developed, and the flowers are borne in racemose inflorescences arising from the axils of long-shoot leaves. In fruit, wings do not develop on the dorsal surface of the bracts of the female flowers. Long shoots do not branch as actively at the top of the crown. …… 3
3. Inflorescences are large, reaching 30 cm in diameter, and form cymose clusters near the branch tips. …… 4
3. Inflorescences are smaller, up to 15 cm in length and width, and are borne successively on the upper parts of long shoots. …… 5
4. Juvenile plants branch profusely from the base. Adult plants have rather many lateral branches. Leaves on the short shoots are obovate, rounded at the apex, and hairy when young. …… Alluaudia procera
4. Juvenile plants do not branch from the base. Adult plants have few lateral branches. Leaves on the short shoots are circular, with a notched apex, and glabrous. …… Alluaudia montagnacii
5. Tree reaching 12 m in height, with a thick trunk and few, erect lateral branches. Leaves on the short shoots are circular, with a notched apex. In female flowers, the wings of the two bracts are almost equal in size. …… Alluaudia ascendens
5. Shrub or small tree reaching 5 m in height, with a slender trunk and numerous, elongated, somewhat spreading lateral branches. Leaves on the short shoots are obovate, with the apex rounded, truncate, or slightly emarginate. In female flowers, the wing of one bract is markedly larger than that of the other. …… Alluaudia humbertii

Alluaudia dumosa_2: cropped a photo by Stephane Philizot, https://www.inaturalist.org/photos/28865282, CC BY-NC

Alluaudia comosa_5: cropped a photo by Norbert Hahn, https://www.inaturalist.org/photos/447263472, CC BY-NC
Alluaudia comosa_6: cropped a photo by CORDENOS Thierry, https://www.inaturalist.org/photos/111983593, CC BY-NC



Alluaudia montagnacii_2: cropped a photo by amantedarmanin, https://www.inaturalist.org/photos/656044045, CC BY
Alluaudia montagnacii_3: cropped a photo by amantedarmanin, https://www.inaturalist.org/photos/656046572, CC BY
Alluaudia montagnacii_4: cropped a photo by amantedarmanin, https://www.inaturalist.org/photos/654333728, CC BY

Rauh, W. (1963). 121 Famille. Didiereacees. In Flore de Madagascar et des Comores: plantes vasculaires, , H. Humbert and J.-F. Leroy, eds. (Museum National D’Histoire Naturelle). CC BY-NC-SA 4.0

Alluaudia humbertii_2: cropped a photo by globalherping, https://www.inaturalist.org/photos/401184181, CC BY-NC
Alluaudia humbertii_3: cropped a photo by dennis-mada, https://www.inaturalist.org/photos/11926595, CC BY-NC
Rauh, W. (1963). 121 Famille. Didiereacees. In Flore de Madagascar et des Comores: plantes vasculaires, , H. Humbert and J.-F. Leroy, eds. (Museum National D’Histoire Naturelle). CC BY-NC-SA 4.0
謝辞 Acknowledgement
本稿執筆にあたり、名古屋市東山動植物園のカナボウノキの栽培株を観察させていただいた。心より感謝申し上げます。
引用文献 References
Bond, W.J., and Silander, J.A. (2007). Springs and wire plants: anachronistic defences against Madagascar’s extinct elephant birds. Proceedings of the Royal Society B: Biological Sciences 274, 1985–1992. https://doi.org/10.1098/RSPB.2007.0414.
Bruyns, P. V., Oliveira-Neto, M., Melo-de-Pinna, G.F., and Klak, C. (2014). Phylogenetic relationships in the Didiereaceae with special reference to subfamily Portulacarioideae. Taxon 63, 1053–1064. https://doi.org/10.12705/635.36.
Carolin, R.C. (1993). Portulacaceae. In The Families and Genera of Vascular Plants. Volume II. Magnoliid, Hamamelid and Caryophyllid Families, K. Kubitzki, J. G. Rohwer, and V. Bittrich, eds. (Springer), pp. 544–555.
Choob, V. (2022). Prophyll in Monocots: The Starting Point of Lateral Shoot Phyllotaxis. Front. Plant Sci. 13, 855146. https://doi.org/10.3389/FPLS.2022.855146/TEXT.
Crowley, B.E., and Godfrey, L.R. (2011). A glance to the past: subfossils, stable isotopes, seed dispersal, and lemur species loss in Southern Madagascar. Am. J. Primatol. 73, 25–37. https://doi.org/10.1002/AJP.20817.
Erbar, C., and Leins, P. (2006). Floral ontogeny and systematic position of the Didiereaceae. Plant Syst. Evol. 261, 165–185.
Kubitzki, K. (1993). Didiereaceae. In The Families and Genera of Vascular Plants. Volume II. Magnoliid, Hamamelid and Caryophyllid Families, K. Kubitzki, J. G. Rohwer, and V. Bittrich, eds. (Springer), pp. 292–295.
Marloth, R. (1913). The Flora of South Africa, Volume I (Darter Bros. & Co.).
Ocampo, G., and Columbus, J.T. (2010). Molecular phylogenetics of suborder Cactineae (Caryophyllales), including insights into photosynthetic diversification and historical biogeography. Am. J. Bot. 97, 1827–1847. https://doi.org/10.3732/AJB.1000227;SUBPAGE:STRING:FULL.
Pax, F. and Hoffmann, K. 1936. Capparidaceae. In: Engler, A. & Prantl, K. (eds.), Die natürlichen Pflanzenfamilien, 2nd ed., Band 17b: 146–223. Leipzig: Wilhelm Engelmann.
Pearson, H.H.W., and Stephens, L. (1912). List of the plants collected in the Percy Sladen Memorial Expedition, 1908-1909, 1910-1911: Portulacaceae. Ann. South African Museum 9, 30–35.
POWO (2026). Plants of the World Online. Facilitated by the Royal Botanic Gardens, Kew. Published on the Internet; https://powo.science.kew.org/. Retrieved 7 July 2026.
Rauh, W. (1956). Morphologische, entwicklungsgeschichtliche, histogenetische und anatomische Untersuchungen an den Sprossen der Didiereaceen. Abhandlungen der Akademie der Wissenschaften und der Literatur, Mathematisch-Naturwissenschaftliche Klasse 1956 (6), 369–540.
Rauh, W. (1960/1961). Weitere Untersuchungen an Didiereaceen. 1. Teil: Beitrag zur Kenntnis der Wuchsformen der Didiereaceen, unter besonderer Berücksichtigung neuer Arten. Sitzungsberichte der Heidelberger Akademie der Wissenschaften, Mathematisch-Naturwissenschaftliche Klasse, Jahrgang 1960/1961, 185–300.
Rauh, W. (1963). 121 Famille. Didiereacees. In Flore de Madagascar et des Comores: plantes vasculaires, , H. Humbert and J.-F. Leroy, eds. (Museum National D’Histoire Naturelle).
Rauh, W. (1983). The morphology and systematic position of the Didiereaceae of Madagascar. Bothalia 14, 839–843. https://doi.org/10.4102/ABC.V14I3/4.1251. Troll, W., and Rauh, W. (1950). Das Erstarkungswachstum krautiger Dikotylen, mit besonderer Berücksichtigung der primären Verdickungsvorgänge. Sitzungsber. Heidelb. Akad. Wiss. Math. Naturwiss. Kl. 1950, 1–86.