
ステート遷移の構造基盤
太陽光は全ての生命にとって重要なエネルギー源です。植物は光合成反応を利用して太陽光のエネルギーを獲得しますが、その際の光の利用効率は常に最適化されています。その最適化のしくみの一つが「ステート遷移」です。葉緑体のチラコイド膜には光化学系IIと光化学系Iと呼ばれる2つの光化学系が直列に並んでおり、この2つの光化学系が光エネルギーから電子伝達を生み出します。この2つの光化学系はエネルギー順位的に直列に並んでいるため、一連の電子移動を効率よく行うためには、2つの光化学系をバランスよく駆動する必要があります。バランスが崩れると電子移動が滞り植物にストレスがかかることになります。太陽光の色や強さは刻一刻と変化しますから、自然界の中ではこのバランスが崩れることはよくあります。そのようなとき、瞬時にバランスを取り戻すしくみがあります。このしくみは、1969年に村田紀夫・元基礎生物学研究所教授により発見され「ステート遷移」として知られてきました。その後、光化学系IIが過度に励起されると集光アンテナLHCIIがリン酸化されて切り離されて光化学系Iに再結合することなどが少しずつわかってきました。
わたしたちは、緑藻クラミドモナス細胞のLHCIIのリン酸化を人工的におこしてステート2と呼ばれる状態を作り出し、光化学系I(PSI)-集光装置I(LHCI)-集光装置II(LHCII)からなる超分子複合体(以下超複合体)を獲得しました。そして数千に及ぶクライオ電子顕微鏡画像を取得した上でコンピュータ画像処理(単粒子解析)により立体構造を解明しました。野生株細胞および“ステート2固定変異株”(pph1;pbcp)より単離された一つ目の超複合体は、片側に2層の”LHCIベルト”を、もう片側に”LHCIハーフベルト”と2つのモバイルLHCII三量体を結合した構造であり解像度2.8Å(1 Åは1mmの一千万分の一)で決定しました(Fig.1)。
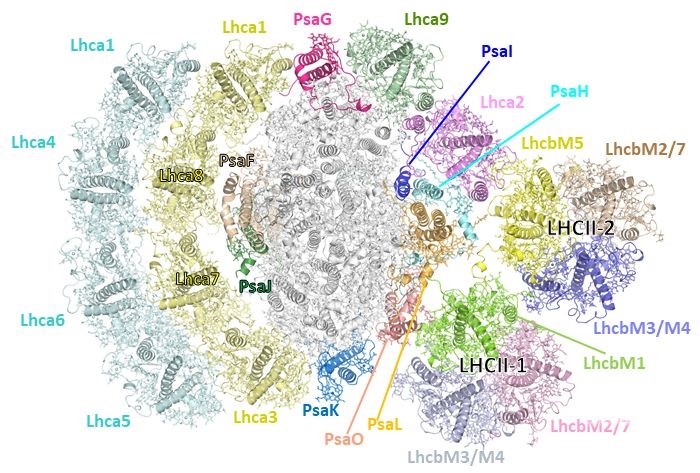
Fig.1. 光化学系Iステート2超複合体(PSI-LHCI-LHCII)構造.葉緑体チラコイド膜鉛直方向ストロマ側から見下ろした全体構造.中央部分(グレイ)が光化学系I(PSI)であり、その周りを以下の集光アンテナが取り囲む構造である。左側に2層の「LHCIベルト」(黄、シアン)が結合し、右側上部には、Lhca2-Lhca9の2分子からなる「LHCIハーフベルト」が結合している。さらに、右側中央から下部にかけて、2つのモバイルLHCII三量体(LHCII-1, LHCII-2)が結合している。
今回3Å以下の高解像度で構造が解明されたことから、光化学系IIとの間で受け渡されるとされるモバイルLHCII三量体の全サブユニットが同定されました。Fig.2aでは右下に示されているLHCII-1では、LhcbM1サブユニット(緑)がPSI(グレイ)と接しており、他にLhcbM2/7とLhcbM3/4サブユニットが同定されました。また、その上に示されているLHCII-2では、LhcbM5サブユニット(黄)がPSIと接しており、他にLhcbM2/7とLhcbM3/4サブユニットが同定されました(Fig.2a)。さらに、LhcbM1のN末端部分の27番目のスレオニン残基(Fig.2b)とLhcbM5のN末端部分の33番目のスレオニン残基(Fig.2c)がリン酸化修飾を受けており、アルギニンなどプラス電荷を帯びるアミノ酸残基で構成されるPSI側の結合部位と安定に相互作用していることもわかりました。これまでの機能解析の結果から、ステート遷移においてモバイルLHCII三量体のN末端部分のリン酸化が重要であることはわかっていましたが、今回それが細部に至るまで立証されたことになります。
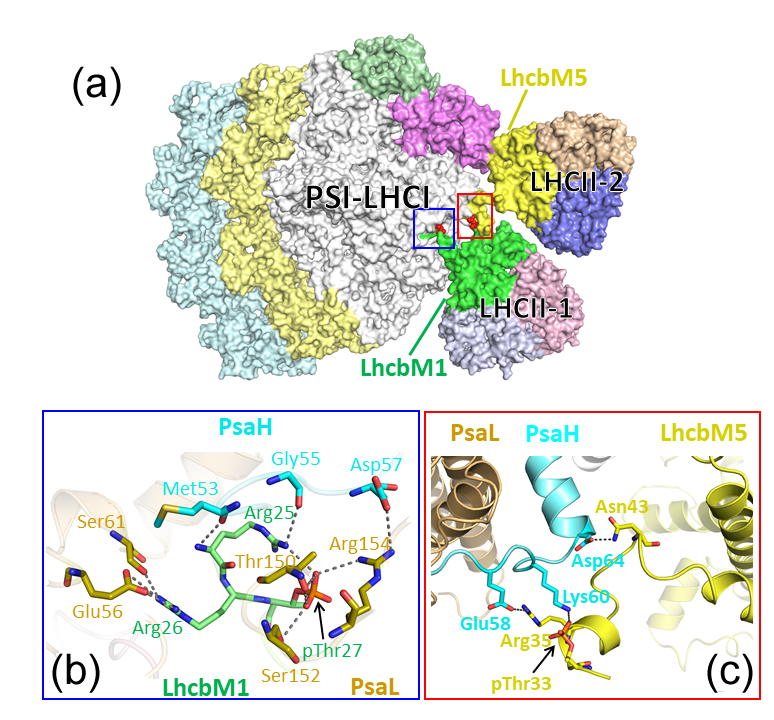
Fig.2. モバイルLHCII三量体結合面の詳細.(a) 光化学系Iステート2超複合体の全体像(膜鉛直方向、ストロマ側から)。LHCII-1はLhcbM1サブユニットのリン酸化された27番目のスレオニン残基 (b) をLHCII-2はLhcbM5サブユニットのリン酸化された33番目のスレオニン残基 (c) をPSI側サブユニットのアルギニン残基等と相互作用させて、ひっかけるように“ゆるく”結合している。
また、各サブユニットの位置、各クロロフィルの位置と配向が決定されたことで、クロロフィル間の励起エネルギー移動効率の計算が行われました。2つのモバイルLHCII三量体はLHCIIハーフベルトと一体化し、互いの間でエネルギーを素早くやりとりする一つのアンテナベルトとして機能していることもわかりました。今回解明された構造は、光合成生物が太陽光エネルギーの成分をいかに効率よく吸収・利用しているのかを理解するために役立ちます。モバイルLHCIIの各サブユニットが同定された意義は大きく、今後はそのサブユニットがPSII-LHCII超複合体のどこからはずれてやってきたのか(あるいは別のところからやってきたのか)研究が加速されるでしょう。
本研究で私たちと共同研究を行ったMei Li博士のグループは、2018年に陸上植物であるトウモロコシのPSIステート2超複合体の構造を決定しています。その構造と今回の緑藻のPSIステート2超複合体の構造を比較すると共通点と相違点が浮かび上がります。陸上植物のPSIには1層のLHCIベルトしかなく、LHCIハーフベルトもありません。そのためか、ステート遷移でも1分子のモバイルLHCII三量体しか結合しません。一方緑藻は2層のLHCIベルトを持っています。さらにLHCIハーフベルトを結合しており、ステート遷移では、まずモバイルLHCII-2が安定化され、さらにモバイルLHCII-1が結合して全体として一つのアンテナベルトを構成していることがわかりました(Fig. 3)。
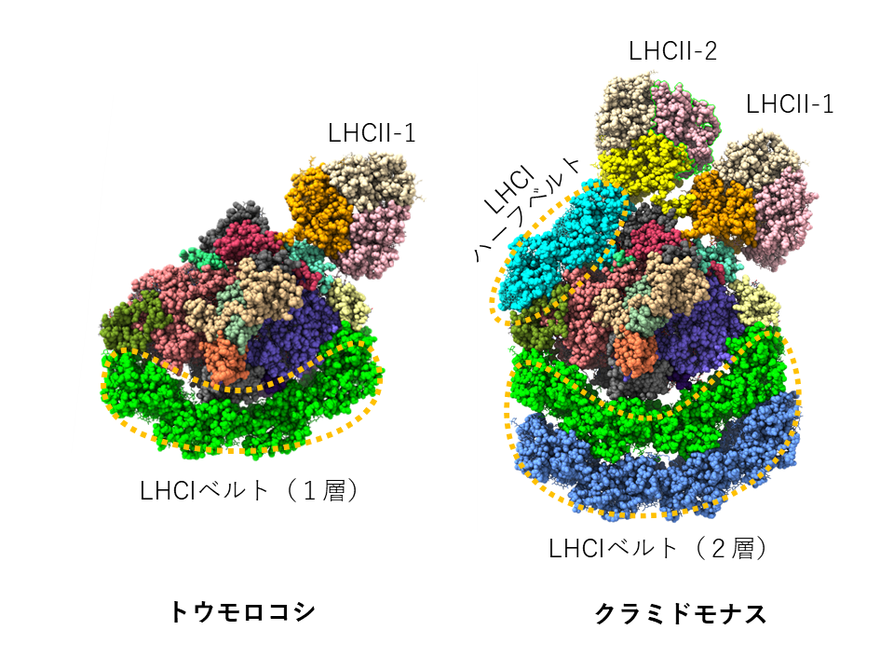
Fig.3. 陸上植物と緑藻のPSIステート2超複合体の集光アンテナの比較.中心に位置するPSIにて光化学反応(光エネルギーが電気化学エネルギーに変換される)が行われるが、そこに光エネルギーを供給する集光アンテナの構造が陸上植物と緑藻では異なる。通常時はモバイルLHCII-1やLHCII-2は結合しておらず、PSIとPSIIの励起バランスを補正する必要が生じたステート2の時のみ、これらのモバイルLHCII三量体は結合する。