
緑藻は二重の強光順化によって守られている
植物(陸上植物や藻類)は、動物のように活発に動くことができないため、自分たちが好きな環境に移動できません。このため、たとえ周囲が高温、低温、乾燥といった過酷な環境になったとしても、その場所で生き残っていく必要があります。そのために、植物は体内の様々な生物反応を時にはダイナミックに、時には微細に調節しながら自らを周囲の環境に馴らしています。植物は光を受けて光合成を行うことでエネルギーを作り出し成長しますが、強すぎる光は植物にとって有害であることが知られています。特に曇り空から雲が去り、急に晴れ間がさした時などは、急激に光の強さが変わるため、強い光に対して迅速に適応しなければ、光合成器官が破壊されてしまいます。光合成器官の破壊は、まさしく植物の生死に直結しています。そうならないように、光合成器官もまた、強い光のもとでは精密に調節されていると考えられています。しかし、実際のところ植物がどのように強い光に適応しているのか、その全体像はよくわかっていません。これまでの研究から、植物に強い光が照射された場合の適応反応として、qEクエンチングと呼ばれる『余分な光エネルギーを消去する』反応が有効であることが分かってきています。しかし、この反応だけで十分なのか、qEクエンチング反応が駆動されるまでの間(植物種によっては数時間かかる)は、どのように強い光をしのいでいるのか、といった点が謎でした。
今回、研究グループは,単細胞緑藻であるクラミドモナスに強い光を当て,どのように強光に適応しているのかを詳しく調べました。強い光の影響を最も受けるのは光合成そのものを行うタンパク質です。研究グループは今回の研究により、強い光の被害を最も受けやすいPSIIと呼ばれるタンパク質複合体に注目し、生理学的・生化学的に分析することで、緑藻が2つの異なる反応を経時的に駆使して強い光に適応することを証明しました。その実態は、強光が照射されてから最初のうちはPSIIタンパク質複合体から『光を集めるアンテナを切り離す』数分間で完了する反応でしのぎ、強い光を当ててから4時間後にはqEクエンチングも導入し『余分な光エネルギーを消去』するというものでした(図1)。

図 1. 緑藻で明らかとなった二重の強光馴化
光合成では通常、まず光を集めるアンテナを使って必要な光を集め、その光エネルギーを利用しています。しかし、先に述べたように、強すぎる光は光合成の反応中心(PSIIタンパク質複合体)を壊す危険性があるため、強光が当たった植物は逆にqEクエンチング反応により光エネルギーを捨てはじめます。緑藻の場合はLHCSRと呼ばれるタンパク質がqEクエンチングの要となっていますが、このタンパク質の合成には少なくとも4時間必要であることが分かっています。しかし、光の変化はとても早いので4時間もの間強い光にさらされてしまうことは、植物(光合成)にとってとても危険です。今回の研究により、この4時間の間は光を集めてもPSIIに渡さないように、光を集めるアンテナをリン酸化修飾することで、一時的にPSIIから切り離すことが分かりました。この一時的な対処は『ステート遷移』と呼ばれており、これまでは光の色(夕日や水中での光)の変化に対応するためのものだと考えられていました。今回の研究から、ステート遷移が、突然の強い光に短期的に対応し、“とりあえず、この場をしのぐ”形で駆動することが分かりました。そして4時間後、たくさん合成されたLHCSRタンパク質を使って“本格的に光エネルギーを消去する”ためにqEクエンチングを駆動します。面白いことに、4時間を超えてさらに強い光にさらされ続けると、今度は、ステート遷移の逆の反応が起こる(一度切り離された光捕集アンテナがふたたびPSIIへと戻ってくる)ことが分かりました(図2)。同時にqEクエンチングの要となるLHCSRタンパク質がPSIIへと結合することが分かりました(図3)。この“逆”のステート遷移反応により、最大限qEクエンチングを活性化させて強光馴化しているのです。

図 2. LHCII の脱リン酸化により明らかになった強光下(HL 条件)での逆ステート遷移(S2→S1)の進行
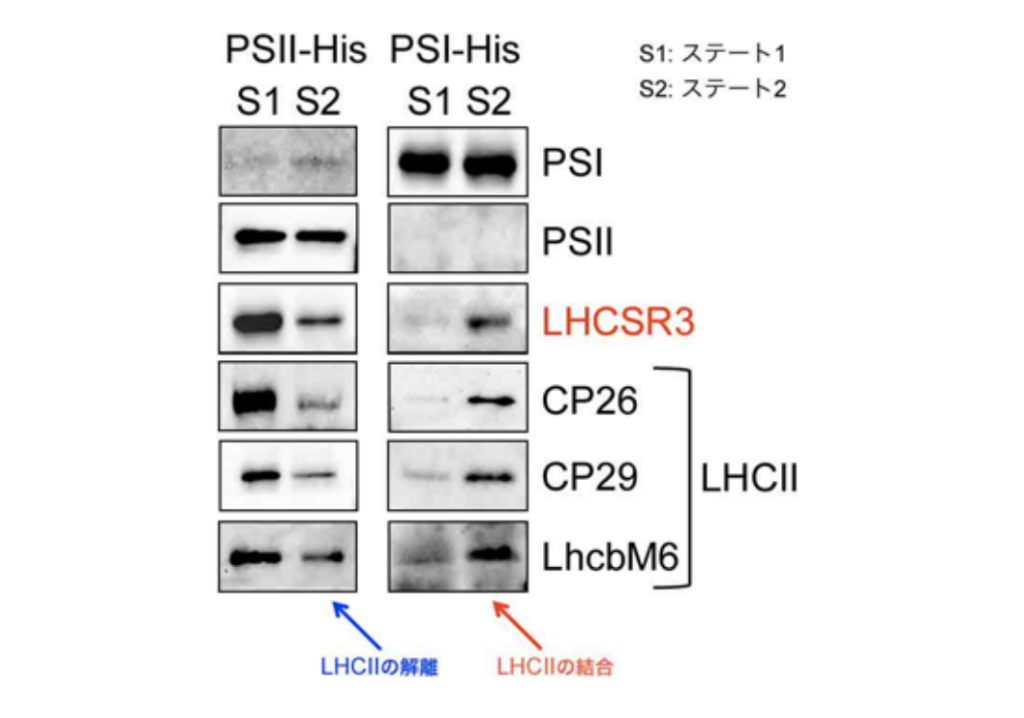
図 3. ニッケルカラムにより精製した光化学系 I(PSI)と光化学系 II(PSII)の成分分析。
ステート遷移の前(S1)と後(S2)では、各光化学系に結合する光のアンテナ(LHCII)や LHCSR3の量が異なる。