
強すぎる光から光化学系をまもるしくみにE3リガーゼが関与
植物細胞の中には、葉緑体という細胞内小器官(オルガネラ)があります。葉緑体の中にはチラコイドと呼ばれる膜でできた袋構造があり、その表面にはたくさんの光合成装置が並んでいます。ここにあたった光のエネルギーを使って電子が流れるとATPができます。電子とATP、そして取り入れた二酸化炭素を固定して糖分を作る一連の反応が『光合成』です。光合成装置は精巧にできているがために、強すぎる光を当てると壊れてしまうという厄介な性質があります。このため、植物は、強い光を浴びたときに、そのエネルギーをわざと逃がすしくみを備えています。この安全装置(qEクエンチング(*1))は、水中で光合成を行う藻類から陸上植物にいたるまでほぼ全ての植物に備わっており、現代光合成研究の中心的なテーマになっています。研究グループは、これまで単細胞植物とも言うべき緑藻クラミドモナスにおいて、葉緑体の中の光化学系IIに結合してqEクエンチングを引き起こすLHCSR3タンパク質に注目してきました(LHCSR3タンパク質の詳細は2013年5月28日のプレスリリース「過剰な光エネルギーを消去する実体,光合成タンパク超複合体を発見」を参照)。そして、光合成とは直接関係ないとされていた青色光受容体の一つフォトトロピンがLHCSR3タンパク質の合成に決定的な役割を果たしていることを国際共同研究で発見しました(2016年9月9日のプレスリリース「青色光受容体が光合成にブレーキをかけることを発見〜青い光が光合成装置を守る〜」を参照)。今回は、さらに青色光シグナルが細胞内で光防御シグナルにいかに変換されるのか、その分子実体に迫りました。
研究グループは、フォトトロピンからの青色光シグナルがいかにLHCSR3遺伝子まで伝わるのかを調べるため、遺伝子ゲノム上からフォトトロピン遺伝子が失われたphot欠失変異株を利用しました。phot欠失変異株は青色光シグナルをLHCSR3遺伝子に伝えることができないためにqEクエンチングがおこらず、クラミドモナスは強い光に耐えることができません。研究グループは、phot欠失変異株のこの性質を逆に利用し、phot欠失変異株に変異を誘発し、たくさんの突然変異株の中から強い光のもとでも生きることができる抑圧変異株を複数取得することに成功しました(Fig.1)。

Fig.1. 抑圧変異株のスクリーニング
野生株プール(WT)、突然変異処理前の phot欠失変異株プール、抑圧変異株プール。縦軸は生育の目安となる自家蛍光強度を示す。赤枠で囲んだ部分が、強光で生存する抑圧変異株。
得られた抑圧変異株のうち3株の解析を進めたところ、ヒトを含め動物・植物に広く分布しているCUL4型E3ユビキチンリガーゼ(*2)のサブユニット変異株ddb1-1、さらに、それと相互作用する因子の変異株det1-1,det1-2であることがわかりました。これらのE3リガーゼ因子は陸上植物では光形態形成において重要な役割を果たすことがわかっており、その変異植物は暗所でも芽生えがおこります。一方、ヒトでは細胞周期や細胞死に関わっているとされ、がん抑制遺伝子や前がん遺伝子の制御に関わっているとの報告もあります。研究グループは、得られた3つの抑圧変異株の解析を進め、DDB1、DET1、CUL4が互いに複合体を形成していること(CUL4-DDB1DET1)、これらの抑圧変異株では、LHCSR3タンパク質ばかりでなく、qEクエンチングを引き起こすもうひとつの重要タンパク質であるLHCSR1タンパク質の遺伝子合成も誘導され(LHCSR1については、2018年3月20日のプレスリリース「植物が強すぎる光エネルギーを逃がす新たな仕組みが見つかる」を参照)、大きな強光耐性能力(NPQ値)が実現されること(Fig.2)、そして、強光条件では抑圧変異株は対照株よりも良好に生育すること(Fig.3)などを明らかにしました。
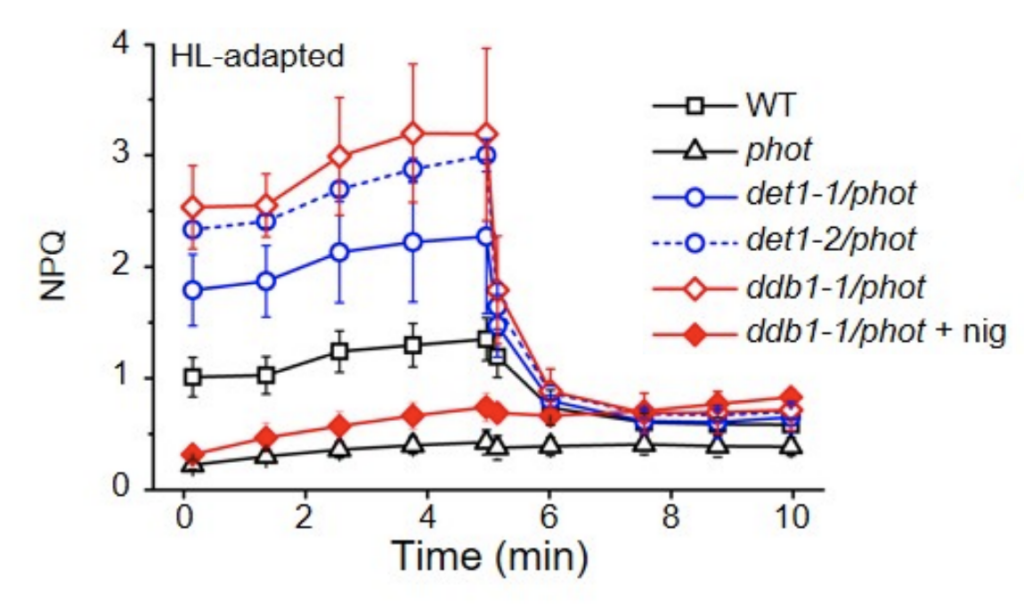
Fig.2. 抑圧変異株の強光耐性能力(NPQ値)
野生株(WT)の NPQ 値が 1.0 前後、phot 欠失変異株の NPQ 値が 0.25 前後であるのに対し、抑圧変異株の NPQ 値は 2~3と高い。

Fig.3. 抑圧変異株の強光条件における生育
強光条件では phot 欠失変異株は育つことができない。また野生株(WT)も生育は制限される。一方、抑圧変異株は、強光条件でも良好な生育を示した。
*1
qEクエンチング:クエンチング(q)とは一般に(火などが)“消える”、(乾きを)“癒やす”などの意味で使われるが、光合成反応においては、光エネルギーを吸収して興奮状態になったクロロフィルが鎮まることを意味する。その際の分子機構の違いにより、qEクエンチング、qTクエンチング、qMクエンチングなどが知られている。qEクエンチングは、光合成装置に電子が流れ葉緑体にエネルギー(E)が与えられたときに生じるフィードバック型のクエンチングのこと。NPQ(ノン・フォトケミカル・クエンチング;光化学反応によらないエネルギー消去反応)とも呼ばれる。
*2
E3ユビキチンリガーゼ:ユビキチン転移酵素。ユビキチンとはあらゆる真核生物に存在する小さなタンパク質。E3ユビキチンリガーゼはユビキチンを特定の標的タンパク質に付加する働きを持つ。数百種類あると言われ、それぞれ標的タンパク質が異なる。”ユビキチン化”を受けた標的タンパク質は、分解を受けたり活性変化したりしてその機能が制御される。