
光化学系II-集光装置超複合体の立体構造を決定
植物細胞の中には葉緑体という細胞内小器官(オルガネラ)があります。葉緑体の中にはチラコイドと呼ばれる生体膜でできた袋構造があり、その表面にはたくさんの膜タンパク質複合体がありエネルギー関連の生理反応を行っています。光化学系I、光化学系IIと呼ばれる二種の膜タンパク質色素超複合体はそのような複合体の仲間であり、太陽光エネルギーを集めて電気化学エネルギーに変換する光合成反応の根幹部分を担っています。光化学系IIは光エネルギーを電気化学エネルギーに変換しますが、その際に水が酸化されて酸素が発生することでも知られています。糖を生産する光合成反応の起点となること、そしてわれわれが呼吸する酸素を生産すること、この2点で光化学系IIは極めて重要な酵素です。光化学系IIは周囲をクロロフィル等の多数の色素を結合した集光装置(light-harvesting complex)で取り囲まれており、それらの集光装置は光化学系IIのために効率よく光エネルギーを集めるという役割を持っています。しかしその集光反応やエネルギー変換反応の詳細を理解するためには、関係するクロロフィル等の色素の位置や配向、周囲のタンパク質アミノ酸残基の情報等が必要で、光化学系IIとその集光装置の構造研究はこれまで盛んに行われてきました。
今世紀に入り、光化学系IIと集光装置それぞれ個別の立体構造はX線結晶構造解析によって原子レベルまで解明が進みました。しかし、光エネルギーがどのように集められ、そしてそれがどのように光化学系IIに届けられるのかその全体のエネルギーの流れを理解するためには、集光装置と光化学系IIが統合された構造(超複合体の構造)を解き明かす必要があります。この超複合体は分子量が100万を超える巨大な複合体となるため、結晶を作製することが困難で研究はなかなか進みませんでした。この状況にブレークスルーを起こしたのがノーベル化学賞(2017年)を受賞したクライオ電子顕微鏡法です。結晶化を経ずともクライオ電子顕微鏡画像の単粒子解析を行うことで超高解像度の立体構造を明らかにすることが可能になりました。中国科学院の柳振峰教授らはこのクライオ電子顕微鏡法をホウレンソウに応用し、陸上植物型光化学系II-集光装置超複合体の立体構造を世界に先駆けて超高解像度で解明しています。
現在地球上の光合成生産の約半分は陸上、残りの半分は水中(海、湖等)で行われていますが、太古の地球上には水中植物(藻類)しか棲息しておらず、それが数億年前に陸上化し、その子孫が現在の陸上植物にあたるとされています。このため、藻類は一般に始原的な光合成の仕組みを備えていると考えられています。これまでの研究から、基本的なしくみは同じものの藻類の方が一般に大きな集光装置を備えていることがわかっています。水中の方が光の強さが弱く大きな集光能力が必要とされているからかもしれません。研究グループは、これまで緑藻を用いて藻類の光合成集光反応におけるさまざまなメカニズムを研究してきました(たとえば2016年9月9日の基礎生物学研究所プレスリリース「青色 光受容体が光合成にブレーキをかけることを発見〜青い光が光合成装置を守る〜」)、今回はこれらの反応が起こる「光化学系II-集光装置超複合体」の立体構造の詳細をクライオ電子顕微鏡法により明らかにしました。
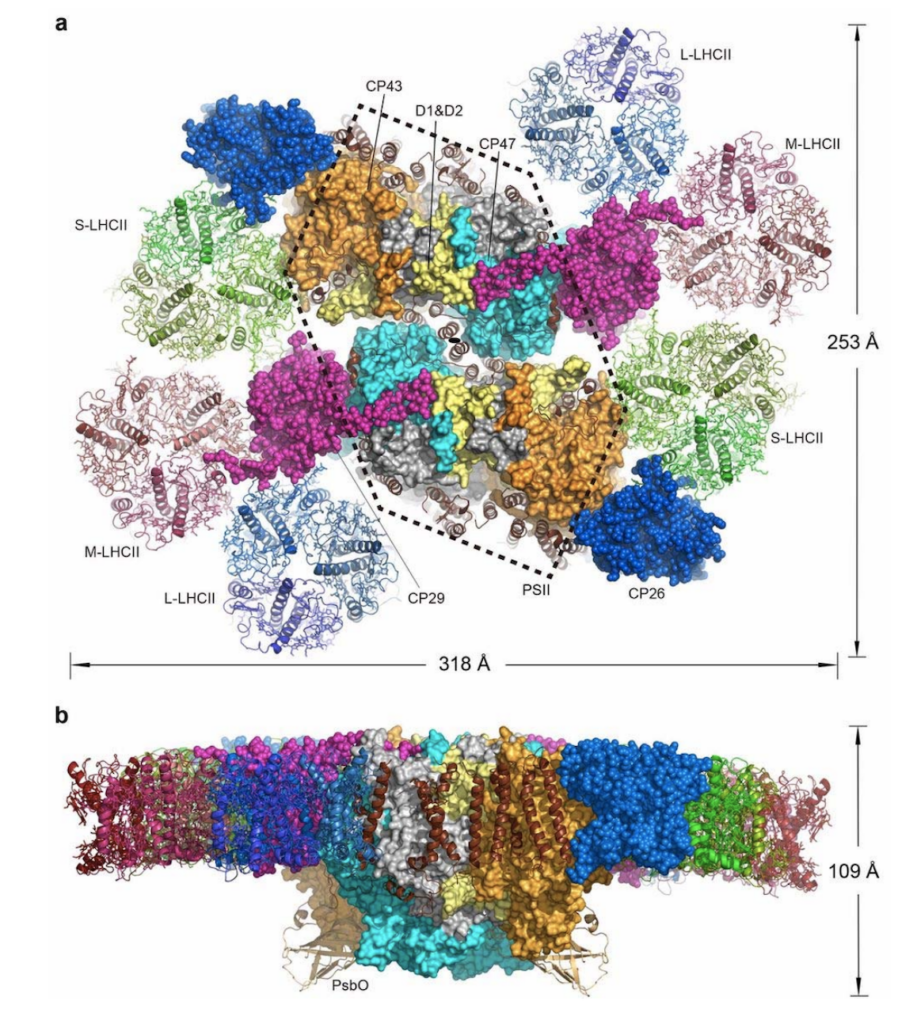
Fig.1.光化学系II-集光装置超複合体C2S2M2L2構造
(a)葉緑体チラコイド膜鉛直方向から見下ろした全景.破線六角形で囲まれた部分が光化学系II(PSII)であり,その両側に集光装置群が結合している.S-LHCII,M-LHCII,L-LHCIIは三量体集光装置。(b)葉緑体チラコイド膜平行方向から見た図.下側の突起部で酸素が発生する。
今回3Åレベルの解像度で構造が解明されたことから、これまで知られていなかった超複合体の内部構造の詳細も明らかになりました。重要な発見の一つは構造脂質の存在です。脂質は従来生体膜成分としてその存在が知られてきましたが、光化学系IIと集光装置をつなぐ“かすがい”の役割をしている糖脂質があることが新たにわかりました(Fig.2)。
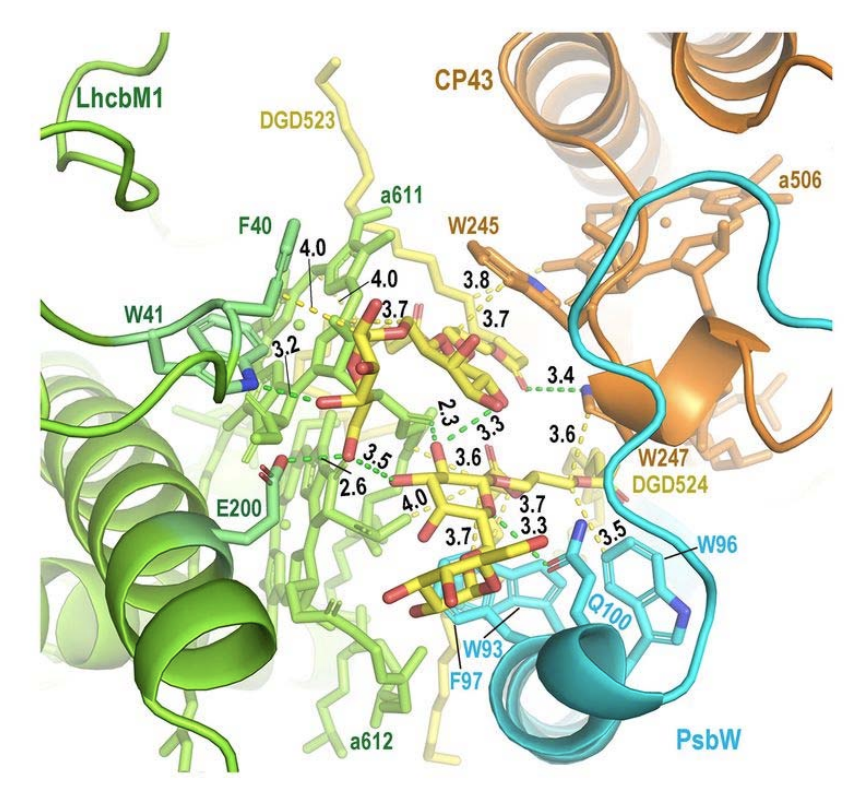
Fig.2. 光化学系 II と集光装置の橋渡しをする糖脂質
光化学系 II(CP43 および PsbW)と三量体集光装置S-LHCII(LhcbM1)は,2 分子のジガラクトシルジアシルグリセロール(DGD523, DGD524)で結合されている。
さらに、CP29と呼ばれる単量体集光装置が2つの三量体集光装置M-LHCII、L-LHCIIと強固なサブ複合体を作っていることもわかりました。今後の詳細な機能解明が待たれます。また、各クロロフィルの位置と配向が決定されたことによって、クロロフィル間の励起エネルギー移動効率の計算も可能となりました。各クロロフィルが光を吸収した後、そのエネルギーがどのような速度でどの程度の効率でどのクロロフィルに移動するのかを予測した結果、緑藻特有の三量体集光装置Lの存在によって全体のエネルギーフローが陸上植物とは異なっていること、そしてこれまで謎に包まれていたクロロフィルZD2がエネルギー伝達に関わっていることなどが明らかになりました(Fig.3)。

Fig.3.クロロフィル間共鳴エネルギー移動
左,時定数10ピコ秒(1ピコ秒は一兆分の1秒)以内におきる共鳴エネルギー移動.主にサブユニット内のクロロフィル間でエネルギー移動が観察される。右, 時定数100 ピコ秒以内におきる共鳴エネルギー移動.集光装置内クロロフィルから光化学系II中心部クロロフィル(Special Pair)までのエネルギー移動ネットワークが観察される。一つ一つの円はクロロフィル分子を示す。星印は本研究で発見されたChlZD2を経由するエネルギー移動のバイパスを示す。