

Pleiostachya pruinosa_1: Cropped a photo by Wayne A. Hughes, https://www.inaturalist.org/photos/149456900, CC BY-NC
Pleiostachya pruinosa-2: Cropped a photo by Kevin Faccenda, https://www.inaturalist.org/photos/550661109, CC BY
Pleiostachya leiostachya-1: Cropped a photo by Marvin López M. , https://www.inaturalist.org/photos/43078880, CC BY-NC
Pleiostachya leiostachya-2: Cropped a photo by Marvin López M. , https://www.inaturalist.org/photos/43026397, CC BY-NC
プレイオスタキア属 Pleiostachya
プレイオスタキア属 Pleiostachya は、中米から南米北部にかけて分布し、3種が知られている (POWO)。P. pruinosaは花序の苞葉にほとんど毛が無いが、P. leiostachyaでは苞葉に軟毛が密生し、葉裏の色も異なる(Hammel 1986)。
The genus Pleiostachya is distributed from Central America to northern South America and comprises three recognized species (POWO). Pleiostachya pruinosa has almost glabrous inflorescence bracts, whereas in P. leiostachya the bracts are densely villous, and the two species also differ in the colour of the abaxial leaf surface (Hammel 1986).

Ischnosiphon petiolatus: cropped a photo by Pierrot F., https://www.inaturalist.org/photos/617327689, CC BY-NC
プレイオスタキア属の特徴 Characteristics of the genus Pleiostachya
プレイオスタキア属は、左右に扁平な花序を形成する点で、姉妹群であるイスクノシフォン属 Ischnosiphonと異なる (Andersson 1998)。
The genus Pleiostachya differs from its sister genus Ischnosiphon in forming laterally flattened inflorescences (Andersson 1998).
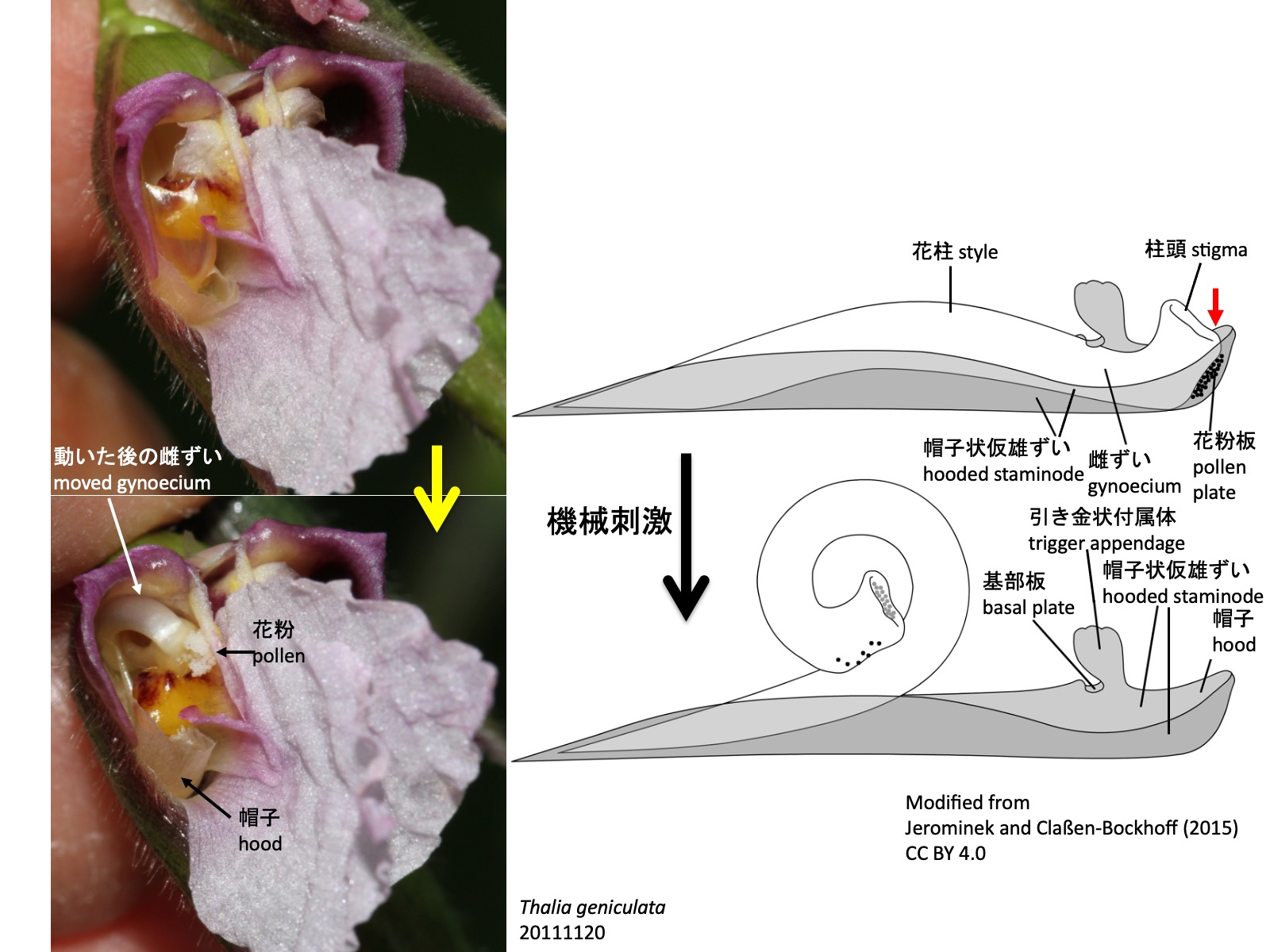
Jerominek, M., and Claßen-Bockhoff, R. (2015). Electrical signals in prayer plants (Marantaceae)? insights into the trigger mechanism of the explosive style movement. PLoS One 10, e0126411. https://doi.org/10.1371/JOURNAL.PONE.0126411. CC BY 4.0.
クズウコン科における花柱運動と二次花粉提示 Style movement and secondary pollen presentation in Marantaceae
クズウコン科の花では、機械刺激によって雌しべの花柱が動き、花粉を二次提示する。蕾の中では、帽子状仮雄ずいの帽子状になった先端部に、雌ずいの先端部、すなわち柱頭の背側と、雄ずい先端部である葯が入り込む。花柱の伸長量が帽子状仮雄ずいの伸長量よりも大きいため、花柱は後方に反るように伸長し、張力が生じる。帽子状仮雄ずいには突起があり、引き金状付属体と、それにつながる基部板を形成している。送粉昆虫が引き金状付属体に触れると、張力を蓄えた花柱を押さえていた基部板が動き、花柱は向軸側へ巻き込む。このとき、花柱は柱頭背側の花粉板に花粉を付けた状態で動くため、花粉が花の前面に提示され、送粉昆虫に付着する。
多くの種では、花粉板と柱頭の間が帽子状仮雄ずいの先端部によってしっかり閉じられているが(模式図の赤色矢印)、Pleiostachya pruinosaではこの密着が緩く、蕾の中で花粉が柱頭に漏れ出し、自家受粉する場合がある (Ley and Claßen-Bockhoff 2012)。このため、温室など送粉昆虫のいない環境でも種子ができやすい。
In flowers of Marantaceae, mechanical stimulation causes the style to move, resulting in secondary pollen presentation. In the bud, the distal hood of the hooded staminode encloses the distal part of the pistil, including the dorsal side of the stigma, together with the distal part of the fertile stamen, namely the anther. Because the style elongates more than the hooded staminode, it grows in a backward-bent position and mechanical tension is generated. The hooded staminode bears a projection that forms the trigger appendage and the connected basal plate. When a pollinating insect touches the trigger appendage, the basal plate that holds the tensioned style in place is displaced, and the style curls adaxially. At this moment, the style moves with pollen attached to the pollen plate on the dorsal side of the stigma; as a result, the pollen is presented at the front of the flower and deposited on the pollinating insect. In many species, the passage between the pollen plate and the stigma is tightly closed by the distal part of the hooded staminode (red arrow in the diagram). In Pleiostachya pruinosa, however, this contact is looser, allowing pollen to enter the stigma in the bud and sometimes resulting in self-pollination (Ley and Claßen-Bockhoff 2012). This may explain why seeds are readily produced even in environments without pollinating insects, such as greenhouses.

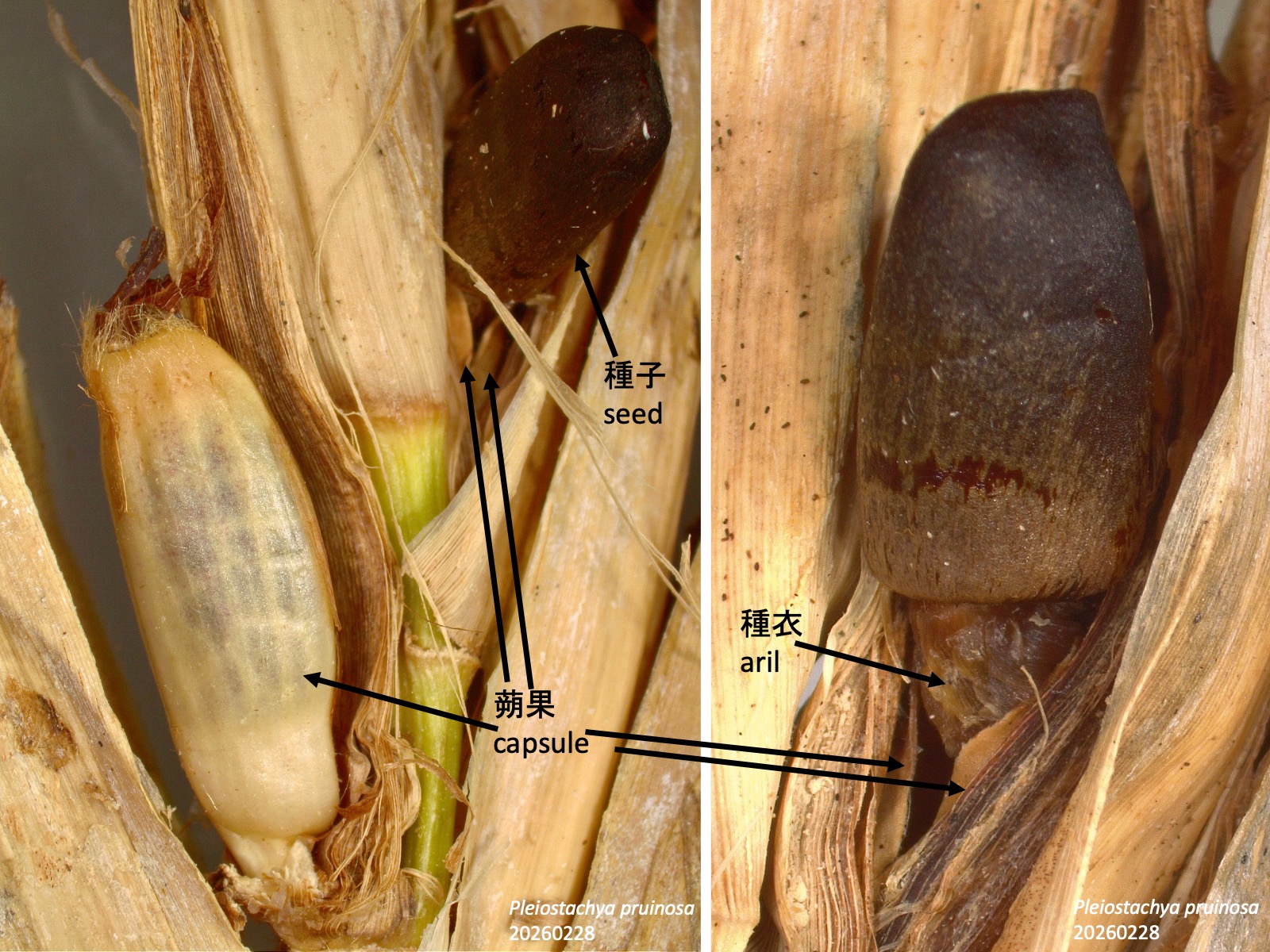
クズウコン科の蒴果から押し出される種子 Seeds protruding from capsules in Marantaceae
クズウコン科では、種子が蒴果内で成熟すると、種衣 arilが発達して膨張し、それに伴って果皮が裂開して、種子が外側へ押し出される (Grootjen 1983)。Pleiostachya pruinosaは、薄茶色に枯れかけた蒴果から、黒い種子が突き出すように露出する (黄色矢印) 。このように露出した種子は、どのように散布されるのだろうか。
In Marantaceae, when the seeds mature within the capsule, the aril develops and expands, causing the fruit wall to dehisce and the seeds to be pushed outward (Grootjen 1983). In Pleiostachya pruinosa, black seeds become exposed as they protrude from pale brown, senescing capsules (yellow arrow). How are these exposed seeds dispersed?

Calathea lutea: cropped a photo by Oscar Johnson, https://www.inaturalist.org/photos/209790037, CC BY-NC-ND
Calathea inocephala: cropped a photo by Vanesa Cruz Jumbo, https://www.inaturalist.org/photos/205659457, CC BY-NC

Pleiostachya leiostachya: Cropped a photo by Eduardo Chacón Madrigal, https://www.inaturalist.org/photos/80935719, CC BY
Goeppertia marantifolia: Cropped a photo by Marco Acuña, https://www.inaturalist.org/photos/324684511, CC BY-NC
クズウコン科の種子散布 Seed dispersal in Marantaceae
クズウコン科Marantaceaeの種には、鳥散布、アリ散布、あるいはその両方による散布が知られている (Horvitz 1991; Horvitz et al. 2002)。コスタリカでは、クズウコン科の数種について、鳥が種子を果序から持ち去る割合、および種子を落葉層の上に置いた場合にアリ類が持ち去る割合を調べる研究が行われた (Horvitz et al. 2002)。
Calathea lutea、Calathea inocephala、Pleiostachya pruinosa、Goeppertia marantifoliaでは、それぞれ異なった形態の種子が蒴果から露出する。C. luteaでは桃色の蒴果から橙色の種皮をもつ種子が、C. inocephalaではオレンジ色の蝋質の蒴果から青色の種子が露出する。P. pruinosaでは薄茶色の蒴果から黒色の種子が露出し、G. marantifoliaでは青色の種子が露出する。なお、上の写真は P. leiostachyaである。
それぞれの種について、鳥が種子を果序から持ち去る割合は、C. luteaでは約80%、C. inocephalaでは約40%、P. pruinosaでは約30%、G. marantifoliaでは約10%であった。一方、種子を落葉層の上に置いて、アリ類が持ち去る割合を調べると、C. luteaではほぼ100%、C. inocephalaではほぼ0%、P. pruinosaでは約40%、G. marantifoliaでは約90%が持ち去られた。ただし、C. inocephalaは異なる調査地で得られたデータである。C. inocephalaは他種に比べて種子が大きく、このためアリ類が運ばなかったのではないかと推測されている。それ以外の種では、果序についているときには鳥に、果序から脱落した後にはアリ類に散布されるが、その依存度は種や生育地によって異なるようである。
In Marantaceae, seed dispersal by birds, by ants, or by both is known in several species (Horvitz 1991; Horvitz et al. 2002). In Costa Rica, a study examined, for several species of Marantaceae, the proportion of seeds removed by birds from infructescences and the proportion of seeds removed by ants when the seeds were placed on the leaf litter (Horvitz et al. 2002).
In Calathea lutea, Calathea inocephala, Pleiostachya pruinosa, and Goeppertia marantifolia, seeds with different morphologies are exposed from the capsules. In C. lutea, seeds with orange arils are exposed from pink capsules, whereas in C. inocephala, blue seeds are exposed from orange, waxy capsules. In P. pruinosa, black seeds are exposed from pale brown capsules, and in G. marantifolia, blue seeds are exposed. The photograph above shows P. leiostachya.
The proportion of seeds removed from infructescences by birds was approximately 80% in C. lutea, 40% in C. inocephala, 30% in P. pruinosa, and 10% in G. marantifolia. In contrast, when seeds were placed on the leaf litter, the proportion removed by ants was nearly 100% in C. lutea, nearly 0% in C. inocephala, approximately 40% in P. pruinosa, and approximately 90% in G. marantifolia. However, the data for C. inocephala were obtained at a different site. Calathea inocephala has larger seeds than the other species, and this may explain why ants did not remove them. In the other species, seeds may be dispersed by birds while they remain attached to the infructescence and by ants after they fall from the infructescence, although the degree of dependence on each disperser appears to vary among species and habitats.
謝辞 Acknowledgements
本稿執筆にあたり、名古屋市東山動植物園の栽培株を観察させていただきました。心より感謝申し上げます。
I am sincerely grateful to the Nagoya Higashiyama Zoo & Botanical Gardens for allowing me to observe cultivated plants for the preparation of this article.
引用文献 References
Andersson, L. (1998) Marantaceae. In K. Kubitzki (ed.) 1998. The Families and Genera of Vascular Plants. Vol. IV. Monocotyledons: Alismatanae and Commelinanae (except Gramineae). Springer. Berlin. Pp. 278-293.
Grootjen, C.J. (1983). Development of ovule and seed in Marantaceae. Acta Bot. Neerl. 32, 69–86.
Hammel, B.E. (1986). The vascular flora of La Selva biological station, Costa Rica Marantaceae. Selbyana 9, 234–242.
Holvitz, C.C. 1991. Light environments, stage structures and dispersal syndromes of Costa Rican Marantaceae. In Huxley, C.R. and Cutler, D.F. eds, Ant-Plant Interactions. Oxford Univ. Press. pp. 463-485.
Holvitz, C.C., Pizo, M.A., y Belllo, B.B., LeCorff, J. and Dirzo, R. 2002. Are plant species that need gaps for recruitment more attractive to seed-dispersing birds and ants than other species? In Levey, D.J, Silva, W.R., and Galetti, M. eds, Seed Dispersal and Frugivory: Ecology, Evolution and Conservation. CABI Publishing, Oxon, UK. pp. 145-159.
Jerominek, M., and Claßen-Bockhoff, R. (2015). Electrical signals in prayer plants (Marantaceae)? insights into the trigger mechanism of the explosive style movement. PLoS One 10, e0126411. https://doi.org/10.1371/JOURNAL.PONE.0126411.
Ley, A.C., and Claßen-Bockhoff, R. (2012). Floral synorganization and its influence on mechanical isolation and autogamy in Marantaceae. Bot. J. Linn.Soc. 168, 300–322. https://doi.org/10.1111/J.1095-8339.2011.01202.X.