
2009年 9月10日
植物の多能性幹細胞の性質を制御する遺伝子の発見、ならびに、陸上植物の祖先についての新仮説の提唱
多能性幹細胞は体を構成するほとんどの種類の細胞を作り出すことのできる細胞です。植物はいろいろな場所に多能性幹細胞を形成し、それぞれが異なった細胞を作ることによって体を形成しています。植物は減数分裂後に開始する1倍体世代と受精後に開始する2倍体世代という二つの世代を持っています。種子を作らない植物では、それぞれに特有の多能性幹細胞が形成され、配偶体(1倍体植物体)と胞子体(2倍体植物体)が形成されます。しかし、配偶体と胞子体の多能性幹細胞がどのような制御のもと形成されるのかは良くわかっていませんでした。基礎生物学研究所生物進化研究部門の岡野陽介研究員、長谷部光泰教授らの研究グループは、科学技術振興機構、金沢大学学際科学実験センターとの共同研究によって、コケ植物のヒメツリガネゴケを材料として、クロマチン修飾を制御するポリコーム2抑制複合体遺伝子(以下ポリコーム遺伝子)を欠失させることにより、配偶体細胞から、受精無しに胞子体多能性幹細胞を形成させることに成功しました。そして、ポリコーム遺伝子が配偶体多能性幹細胞と胞子体多能性幹細胞にどちらになるかの決定に関わっていることを明らかにしました。今回の発見により、異なった種類の多能性幹細胞を形成する分子機構の一端が明らかになったことに加え、受精と幹細胞形成との関係の研究が進展するとともに、将来的には、受精無しで胞子体多能性幹細胞を形成させることが可能となり、植物の育種研究などへの応用が期待されます。さらに、ポリコーム遺伝子を欠失させることによってできた胞子体は絶滅した前維管束植物類に類似しており、これをもとに従来、うまく説明ができなかった陸上植物の初期進化について新しい仮説を提唱しました。この成果は、米国科学アカデミー紀要電子版にて9月10日(日本時間)に発表されました。
[研究の背景]
植物は発生成長過程において、多様な多能性幹細胞を形成します。例えば、花の咲く植物では、2倍体である胞子体の茎や根の先端に多能性幹細胞を形成し、生涯にわたって成長し続けます。一方、種子を作らない植物では、1倍体世代と2倍体世代の2つの世代の両方に多能性幹細胞を形成します(図1)。1倍体世代の多能性幹細胞(配偶体多能性幹細胞)は減数分裂後に形成され配偶体を、2倍体世代の多能性幹細胞(胞子体多能性幹細胞)は受精後に形成され胞子体を、それぞれ作り出します。ところが、高糖濃度培地での培養などのストレスを与えると、配偶体の細胞が受精無しで胞子体多能性幹細胞に変化して胞子体を形成する現象(アポガミー)が知られていました。この場合、できあがった胞子体は受精無しで形成されるため、1倍体ですが、胞子体の発生プログラムが進行します。このことから、胞子体多能性幹細胞形成を制御するようなメインスイッチが存在するのではないかと推定されていましたが、実体は不明でした。
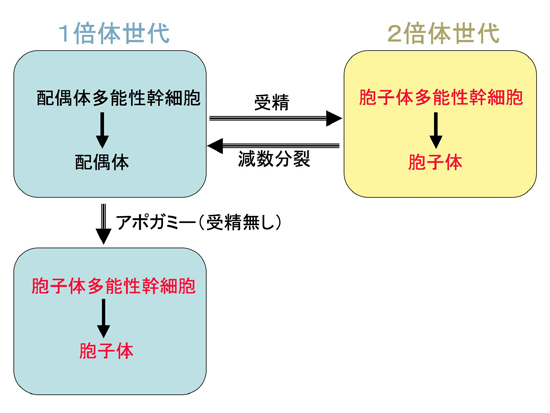
図1:植物の世代交代とアポガミー
植物は1倍体世代に配偶体多能性幹細胞を持ちし、配偶体が形成される。2倍体世代には胞子体多能性幹細胞を持ち、胞子体が形成される。1倍体世代と2倍体世代は受精と減数分裂によって行き来する。一方、アポガミーと呼ばれる現象がおこると、配偶体(1倍体)細胞が受精無しに胞子体多能性幹細胞にリプログラムされ、1倍体の胞子体を形成する。
地球が誕生した後、約5億年前に最初の植物が上陸し、地上は緑に覆われたと推定されています。しかし、初期の陸上植物がどのような形をしていたのかはよくわかっていません。この時代から産出される陸上植物胞子体の大化石(形態が良く保存されていて体の様子が良くわかる化石)はほとんど全て、枝分かれをした体をしていましたが、現在生きている植物には似たものがありません。これら化石植物の中で最も単純な構造をしているのが前維管束植物です(図2)。単純とはいえ、現在生きているコケ植物よりも複雑な形態をしています。従って、化石は無いものの、コケ植物のように枝分かれしない単純な胞子体を持った植物が陸上植物の祖先だったのではないかと考えられてきました(図2)。しかし、前維管束植物と同じ、あるいはそれよりも前の時代にはコケ植物に類似した化石が産出しないことが、この仮説の弱点であり、陸上植物の共通の祖先がどのような形をしていたかを推定する上で大きな問題でした。
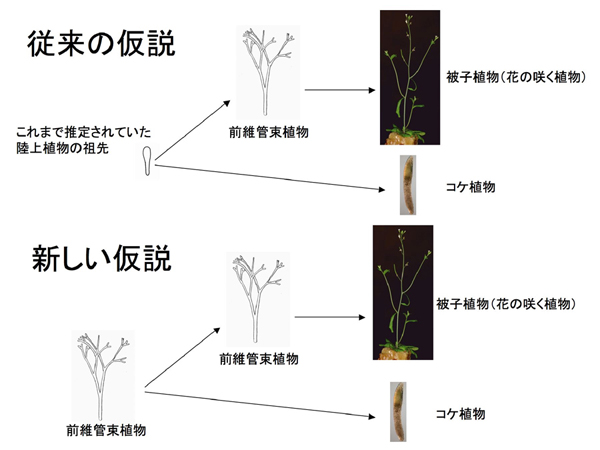
図2:陸上植物の胞子体体制の進化仮説
従来の仮説(上図)では、枝分かれしない、現生コケ植物に似た胞子体を陸上植物の祖先であると仮定していた。そして、現生のコケ植物は祖先からあまり形を変えずに進化したと考えられている(通常我々が目にするコケの体は配偶体で、胞子体は配偶体の先端に半寄生している)。しかし、それを支持するような形態を持つ化石は見つかっていない。また、被子植物などはたくさんの化石が発掘される前維管束植物を経て進化したと考えられている。一方、本研究によって、現生コケ植物ヒメツリガネゴケのポリコーム2抑制複合体遺伝子を欠失させると前維管束植物に類似した胞子体を形成することがわかったことから、コケ植物にも枝分かれする胞子体を作る能力があることが示された。このことから、陸上植物の共通祖先は前維管束植物であり、現生のコケ植物は退化によって進化したという可能性が示唆された。この仮説だと、コケ植物の化石が見つからず、前維管束植物の化石がたくさん産出することに良く合致する。
[研究の成果]
基礎生物学研究所生物進化研究部門の岡野陽介研究員、長谷部光泰教授らの研究グループは、科学技術振興機構、金沢大学学際科学実験センターとの共同研究によって、コケ植物セン類のヒメツリガネゴケを材料として、ヒストン(染色体の中でDNAと結合しているタンパク質)の状態を制御するポリコーム遺伝子(Physcomitrella patens CURLY LEAF遺伝子)を欠失させると配偶体の一部から、受精しないのに、胞子体が形成されるアポガミーと呼ばれる現象が引き起こされることを発見しました。
ヒメツリガネゴケは、配偶体多能性幹細胞である原糸体頂端細胞を形成し、次々と原糸体細胞を作り出し、糸状に生育していきます(図3)。分化した原糸体細胞は、一定の時期になると発生プログラムが変換(リプログラム)され、原糸体頂端細胞を形成します(図3)。
ポリコーム遺伝子はショウジョウバエにおいて初めて発見された遺伝子で、動物、植物に広く保存されており、細胞状態を記憶する働きをしています。具体的には、DNAと結合しているヒストンタンパク質を化学修飾し、修飾を受けた領域のDNA上に位置する遺伝子を働かなくします。ポリコーム2抑制複合体はDNAのいろいろな部分のヒストンに同時に結合することによって、たくさんの遺伝子を同時に制御できると考えられています。
岡野研究員、長谷部教授らのグループは、同グループが以前に解読したヒメツリガネゴケのゲノム情報を利用して、遺伝子ターゲティング法を用いて、ヒメツリガネゴケポリコーム遺伝子の一つであるPhyscomitrella patens CURLY LEAF(PpCLF)遺伝子を欠失するような突然変異体を作出しました。すると、原糸体細胞から、原糸体頂端細胞ではなく、受精後に形成されるはずの胞子体頂端細胞が形成されること、すなわち、アポガミーが引き起こされることがわかりました(図3)。また、PpCLFタンパク質は配偶体の原糸体細胞、精子、卵細胞など配偶体全体で発現し、受精するとその発現が消失することを明らかにしました。このことから、PpCLF遺伝子は、配偶体において、胞子体幹細胞が形成されることを抑制する働きをしていることがわかりました。なお、胞子体において、同遺伝子が生殖器官(胞子嚢)形成を誘導する働きを持つことも明らかになりました。
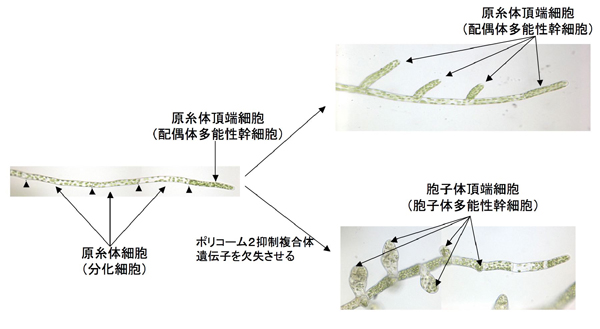
図3:ヒメツリガネゴケの原糸体
先端の原糸体頂端細胞(配偶体多能性幹細胞)が分裂して、原糸体細胞を作り出す。矢尻は細胞の区切りを示す。分化した原糸体細胞は、通常、しばらくするとリプログラムして、原糸体頂端細胞(多能性幹細胞)を形成する。しかし、ポリコーム2抑制複合体遺伝子を欠失させると、胞子体頂端細胞(胞子体多能性幹細胞)が形成されるようになる。
さらに、アポガミーによって形成された胞子体は通常の胞子体と異なり、枝分かれするような胞子体を形成することがわかりました(図4)。しかも、その形態が絶滅した前維管束植物に良く似ていました(図4)。これまで、コケ植物が枝分かれする胞子体を形成できないことから、その単純さゆえ、陸上植物の祖先に最も近い形態を持っているのではないかと考えられてきたのですが、本研究によって、コケ植物も枝分かれする胞子体を形成できることがわかりました。このことから、従来の仮説に加え、陸上植物の共通祖先はたくさんの化石が発掘されている前維管束植物であり、現生のコケ植物は退化によって進化したのだという可能性も検討すべきであることがわかりました(図2)。この新しい仮説だと、従来、祖先と目された現生コケ植物に似た化石が植物が陸上化した年代から産出せず、その後に進化したと想定された前維管束植物の化石ばかりが多数産出するという矛盾が解決できます。

図4:ヒメツリガネゴケポリコーム2抑制複合体遺伝子欠失突然変異体と前維管束植物の比較
ヒメツリガネゴケのポリコーム2抑制複合体遺伝子欠失突然変異体が形成した枝分かれする胞子体(左図)は、絶滅した前維管束植物(右図)によく似ていた。
[今後の展開]
単細胞である卵が、受精後、どのような仕組みで多能性幹細胞を形成するかはほとんどわかっていません。受精後の幹細胞形成にポリコーム遺伝子が関与しているという今回の研究結果は、今後、受精後、多能性幹細胞が形成させるまでの分子機構を解明する足がかりになることが期待されます。また、今回の受精無しで多能性幹細胞形成、胞子体発生過程が進行するという発見を契機に、受精無しで発生過程を進行させる研究がすすみ、将来的には、受精を経ない、すなわち交配しないで作物の種子を大量に作るような技術、あるいは、卵以外のいろいろな細胞から多能性幹細胞を作り出す技術の開発に応用できる可能性があります。また、ヒメツリガネゴケのポリコーム遺伝子を破壊することによって形成される枝分かれした胞子体と前維管束植物化石(図4)との形態的類似点、相違点をより詳細に研究することによって、本研究によって示唆された陸上植物の進化仮説(図2)の妥当性を検討できるようになると考えられます。
[発表雑誌]
Proceedings of the National Academy of Sciences United States of America
米国科学アカデミー紀要 電子版
(9月9日付けでオンライン先行発表)
論文タイトル:
"A polycomb repressive complex 2 gene regulates apogamy and gives evolutionary insights into early land plant evolution
著者:Yosuke Okano, Naoki Aono, Yuji Hiwatashi, Takashi Murata, Tomoaki Nishiyama, Takaaki Ishikawa, Minoru Kubo, and Mitsuyasu Hasebe
[研究グループ]
本研究は、基礎生物学研究所生物進化研究部門(岡野陽介、青野直樹、日渡祐二、村田隆、石川貴章、長谷部光泰)、科学技術振興機構ERATO長谷部分化全能性進化プロジェクト(久保稔、西山智明、長谷部)、金沢大学学際科学実験センター(西山)からなる共同研究チームによる成果です。
[研究サポート]
本研究は、科学技術振興機構、文部科学省科学研究費補助金のサポートを受けて実施されました。
[本件に関するお問い合わせ先]
基礎生物学研究所 生物進化研究部門
教授:長谷部光泰 (ハセベ ミツヤス)
Tel: 0564-55-7546(研究室)
E-mail: mhasebe@nibb.ac.jp
URL: http://www.nibb.ac.jp/evodevo/
[報道担当]
基礎生物学研究所 広報国際連携室
倉田 智子
Tel: 0564-55-7628
E-mail: press@nibb.ac.jp