
![]()
遺伝子のオン・オフを調節するクロマチン構造の謎を解く
細胞が個体を作るための多様性を生み出すためには、決まった時期に適当な遺伝子をオン、あるいはオフにして細胞を特徴づける遺伝子の発現状態を獲得し、また時にはその状態を大きく変化させることで、異なる性質の細胞を生み出す事が重要になります。この際、DNAの一次配列の変化では説明できない、エピジェネティクスと呼ばれる現象が重要な働きをしていると考えられています。私達の研究室では、クロマチンと呼ばれる、DNAとタンパク質の複合体のダイナミックな変化を分子のレベルで解析することで、このエピジェネティクスと呼ばれる生命現象の解明を目的としています。具体的には、クロマチン研究の優れたモデル生物として知られる分裂酵母(Schizosaccharomyces pombe)、ヒト培養細胞、また原生動物である繊毛虫類テトラヒメナを用いて、以下の4つの研究項目を中心に研究を進めています。
1.ヘテロクロマチン構造の形成と維持のメカニズムの解明
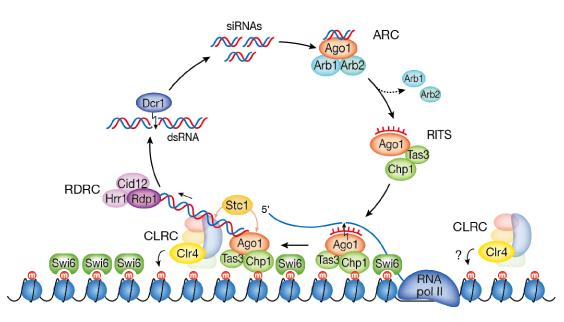 真核細胞の染色体には、高度に凝集したヘテロクロマチンと呼ばれる構造が存在しています。この高次のクロマチン構造は、転位因子や反復配列など、ホストにとって不要な配列を抑制するだけでなく、、セントロメアやテロメアなど染色体の機能ドメインの構築、さらにエピジェネティックな遺伝子発現の制御にも重要な役割を果たしています。この高次のクロマチン構造を形成する分子メカニズムが明らかになれば、複雑なエピジェネティック現象の理解につながると考えられます。また近年の解析から、このヘテロクロマチンの形成過程に、二本鎖のRNAによって引き起こされるRNAサイレンシングの関与が明らかにされてきましたが、その詳細にはまだまだ数多くの謎が残されています。私達の研究室では、主に分裂酵母を材料にして、ヘテロクロマチン構造の形成メカニズムの解明に取り組んでいます。(図:分裂酵母におけるヘテロクロマチン構造形成のモデル)
真核細胞の染色体には、高度に凝集したヘテロクロマチンと呼ばれる構造が存在しています。この高次のクロマチン構造は、転位因子や反復配列など、ホストにとって不要な配列を抑制するだけでなく、、セントロメアやテロメアなど染色体の機能ドメインの構築、さらにエピジェネティックな遺伝子発現の制御にも重要な役割を果たしています。この高次のクロマチン構造を形成する分子メカニズムが明らかになれば、複雑なエピジェネティック現象の理解につながると考えられます。また近年の解析から、このヘテロクロマチンの形成過程に、二本鎖のRNAによって引き起こされるRNAサイレンシングの関与が明らかにされてきましたが、その詳細にはまだまだ数多くの謎が残されています。私達の研究室では、主に分裂酵母を材料にして、ヘテロクロマチン構造の形成メカニズムの解明に取り組んでいます。(図:分裂酵母におけるヘテロクロマチン構造形成のモデル)
2.ヒストンのメチル化を制御する酵素の機能解析
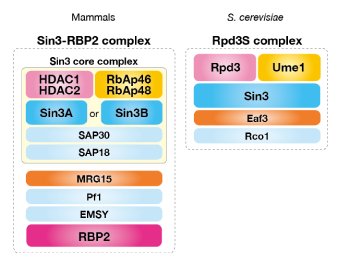 クロマチンの基本単位であるヌクレオソームは、ヒストンと呼ばれる進化的に非常に良く保存された塩基性タンパク質の八量体に、DNAが巻き付く構造をしています。クロマチンの大きな構造変化や、遺伝子の発現調節をもたらすためには、このヌクレオソームの構造を変化させる必要があります。ヒストンのアミノ末端側に飛び出したテイルと呼ばれる領域は、アセチル化、メチル化、リン酸化など、様々な翻訳後修飾を受け、この修飾がエピジェネティックな遺伝子発現制御に重要な役割を果たすことが明らかにされてきました。特にヒストンのメチル化は、安定なエピジェネティックマークとして細胞記憶として働いていることが示唆されています。私達の研究室では、ヒストンのメチル化修飾を触媒するメチル化酵素、脱メチル化酵素とそれらの複合体の機能解析を進めています。(図:脱メチル化酵素RBP2を含む複合体の模式図)
クロマチンの基本単位であるヌクレオソームは、ヒストンと呼ばれる進化的に非常に良く保存された塩基性タンパク質の八量体に、DNAが巻き付く構造をしています。クロマチンの大きな構造変化や、遺伝子の発現調節をもたらすためには、このヌクレオソームの構造を変化させる必要があります。ヒストンのアミノ末端側に飛び出したテイルと呼ばれる領域は、アセチル化、メチル化、リン酸化など、様々な翻訳後修飾を受け、この修飾がエピジェネティックな遺伝子発現制御に重要な役割を果たすことが明らかにされてきました。特にヒストンのメチル化は、安定なエピジェネティックマークとして細胞記憶として働いていることが示唆されています。私達の研究室では、ヒストンのメチル化修飾を触媒するメチル化酵素、脱メチル化酵素とそれらの複合体の機能解析を進めています。(図:脱メチル化酵素RBP2を含む複合体の模式図)
3.クロモドメインタンパク質の機能解析
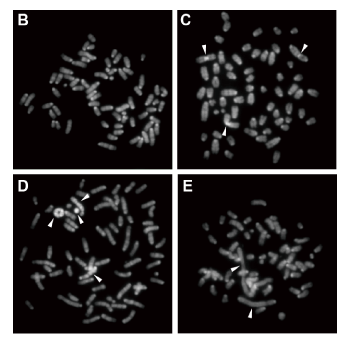 クロモドメインは、クロマチンの構造変化に関わる多くのタンパク質に見いだされる、進化的に保存されたモチーフ構造です。ヘテロクロマチンタンパク質HP1の研究から、クロモドメインがメチル化されたヒストンを特異的に認識して結合する、結合モジュールであることが明らかにされました。ヒストンのメチル化修飾が持つ生物学的な意味を理解するには、その修飾がどのように他のタンパク質に認識されるかを解析することが重要になると考えられます。私達の研究室では、HP1を含む未だ機能が明らかにされていないクロモドメインタンパク質の機能解析に取り組んでいます。また、ヘテロクロマチンタンパク質HP1が、発生や分化における大きなクロマチン構造の変化に果たす、機能的な役割の解明も進めています。(図:HP1のリン酸化欠如によって引き起こされる染色体異常)
クロモドメインは、クロマチンの構造変化に関わる多くのタンパク質に見いだされる、進化的に保存されたモチーフ構造です。ヘテロクロマチンタンパク質HP1の研究から、クロモドメインがメチル化されたヒストンを特異的に認識して結合する、結合モジュールであることが明らかにされました。ヒストンのメチル化修飾が持つ生物学的な意味を理解するには、その修飾がどのように他のタンパク質に認識されるかを解析することが重要になると考えられます。私達の研究室では、HP1を含む未だ機能が明らかにされていないクロモドメインタンパク質の機能解析に取り組んでいます。また、ヘテロクロマチンタンパク質HP1が、発生や分化における大きなクロマチン構造の変化に果たす、機能的な役割の解明も進めています。(図:HP1のリン酸化欠如によって引き起こされる染色体異常)
4.テトラヒメナにおける大規模なゲノム再編成
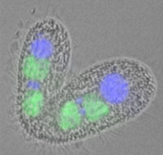 単細胞真核生物であるテトラヒメナは古くからモデル生物として利用され、テロメラーゼ、リボザイム、ヒストンアセチル化酵素などの発見に寄与してきました。テトラヒメナは体細胞核(大核)と生殖核(小核)という二種類の核を持ち、有性生殖時に、小核を元にして大規模なゲノムの再編成が起こって新しい大核が形成されます。興味深いことに、RNAサイレンシングや進化的に保存されたクロモドメインを有するタンパク質が、このゲノム再編成に寄与することが明らかにされてきました。私たちの研究室では、このゲノム再編成の分子機構を解析し、ヘテロクロマチンの進化的意義の解明に取り組んでいます。(図:接合中のテトラヒメナ。青:親大核、緑:新しくできた大核原基)
単細胞真核生物であるテトラヒメナは古くからモデル生物として利用され、テロメラーゼ、リボザイム、ヒストンアセチル化酵素などの発見に寄与してきました。テトラヒメナは体細胞核(大核)と生殖核(小核)という二種類の核を持ち、有性生殖時に、小核を元にして大規模なゲノムの再編成が起こって新しい大核が形成されます。興味深いことに、RNAサイレンシングや進化的に保存されたクロモドメインを有するタンパク質が、このゲノム再編成に寄与することが明らかにされてきました。私たちの研究室では、このゲノム再編成の分子機構を解析し、ヘテロクロマチンの進化的意義の解明に取り組んでいます。(図:接合中のテトラヒメナ。青:親大核、緑:新しくできた大核原基)