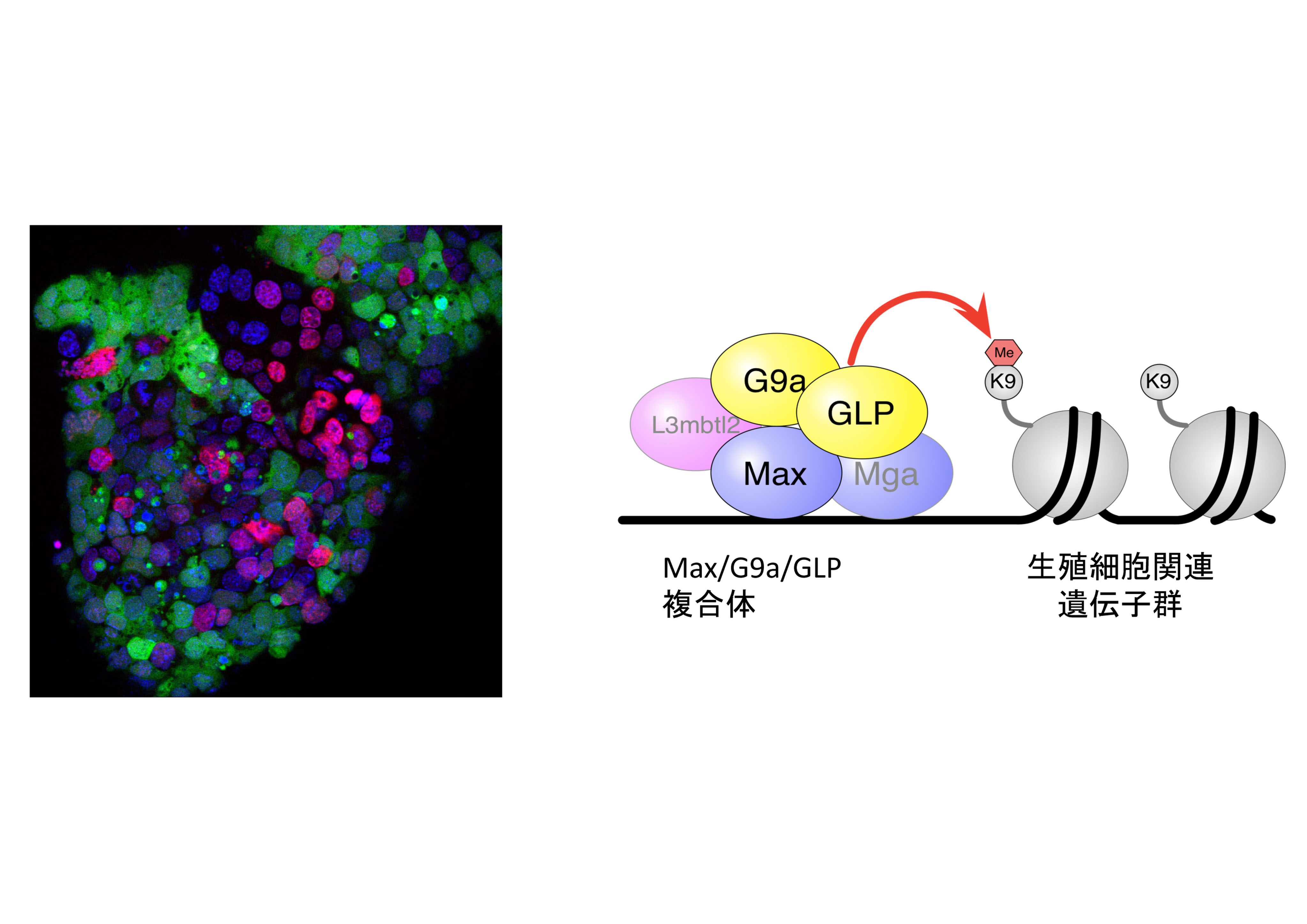
マウス胚発生過程では、多能性幹細胞より分化したエピブラストから始原生殖細胞(PGC)の形成が起こり、また特定の培養条件下などでPGCは多能性幹細胞に比較的容易に変換しうるが、両者は異なる分化能をもつ。本研究では、PGCと多能性幹細胞の違いを生み出している遺伝子ネットワークをマウスで明らかにし、その進化上の普遍性をいくつかのモデル生物を使って解明することを目的とする。本研究代表者らは、これまでの研究で、Maxなどの転写因子がヒストンH3K9のメチル化を介して、ES細胞がPGCへ直接変化することを阻害していることを明らかにした。一方、ニワトリやゼブラフィッシュでは母性決定因子の働きでPGC形成が起こると考えられているが、これら生物の初期胚細胞からES細胞と類似した細胞が得られることから、多能性幹細胞からPGCが形成される可能性がある。さらにプラナリアは、多能性幹細胞であるネオブラストからPGCが形成される。そこでマウス胚およびニワトリ胚、ゼブラフィッシュ胚やプラナリアでも、Max等が多能性幹細胞でのPGC特異的遺伝子の発現抑制に働いているかをRNAiにより明らかにする。