
1.重力屈性の分子メカニズムを探る

図1 シロイヌナズナ花茎の重力屈性
野生型のシロイヌナズナの花茎を水平に倒すことで重力刺激を与え始め(0min)、30分後から100分間10分毎に写真を撮り重ねた。右はある重力屈性異常変異体について同様に写真を撮り重ねたもの。植物とCCDカメラを暗箱内に入れ、写真を撮るときにだけ植物が感じないとされる850nm以上の光を照射して撮影してあるので、光の影響はないと考えてよい。
生き物の多くは、重力方向を感知する能力を持っています。例えば私達は目隠しをされても、どちらが上か下か簡単に分ります。植物もまた重力を感知し、茎は上を、根は下を向いて伸びます。私達はシロイヌナズナを用いて、重力屈性に異常を示す変異体を多数単離し、原因遺伝子の特定とその機能解析を通して重力屈性の分子機構の解明を目指しています。
研究動画
2.重力受容のメカニズム
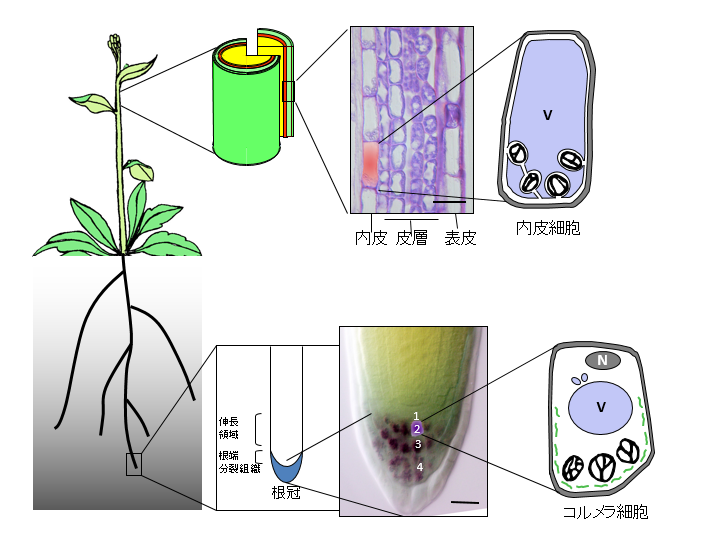
図2 シロイヌナズナの花茎と根における重力感受細胞
シロイヌナズナ花茎の組織構造は、通常外側から1層の表皮、約3層の皮層、1層の内皮が同心円柱状に配置し、さらにその内側に維管束を含む組織がある。内皮細胞ではアミロプラストが重力方向に観察される。内皮細胞は大きく発達した液胞によりその体積のほとんどを占められており、アミロプラストは周囲を液胞膜に取り囲まれているように見える。根の根冠には層状のコルメラ細胞があり、アミロプラストを含んでいる。コルメラ細胞の液胞は比較的小さい。
植物は重力方向の変化をどのようにして感受するのでしょうか?重力屈性をおこなう器官には、澱粉を蓄積した色素体(アミロプラスト)を持つ細胞が観察されます。アミロプラストは周囲の細胞質に比べて比重が高いので、重力の方向に沈む性質があります。従って、器官が傾くと、アミロプラストが重力方向に移動することで重力の方向を感じると考えられています。植物一般に、根の先端の根冠のコルメラと呼ばれる細胞層があり、コルメラ細胞内に重力方向に沈むアミロプラストが観察されます。様々な実験事実から、コルメラ細胞が根の重力感受部位であることが示されています。一方、シロイヌナズナの花茎には、円柱状に並んだ一層の内皮細胞層があり、この内皮細胞中に重力方向に沈むアミロプラストが観察されます。花茎の重力屈性能を完全に失ったsgr1(shoot gravitropism 1), sgr7変異体では、内皮細胞層を欠失しており、このことは内皮細胞が茎の重力感受細胞であることを示しています。
研究動画
3.重力感受の場を視る
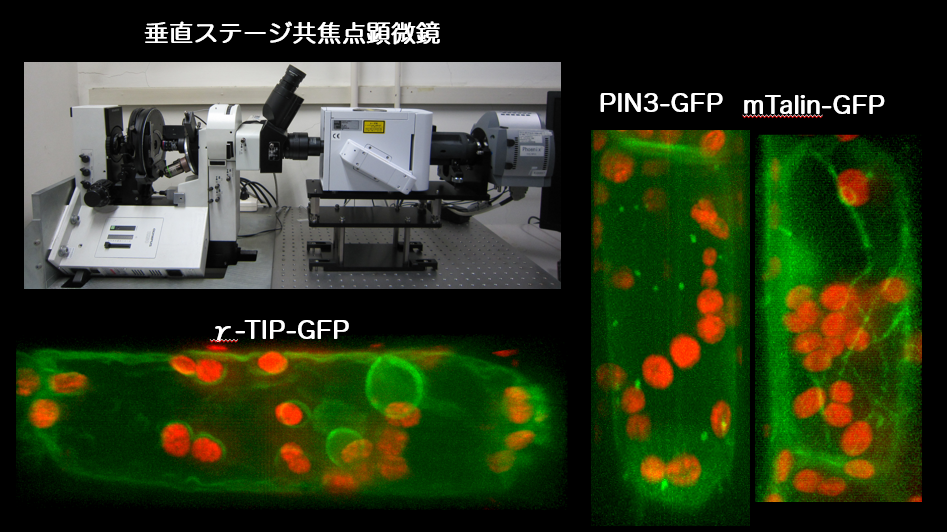
図3 垂直ステージ共焦点顕微鏡を用いた内皮細胞の観察。重力方向を保持して撮影した内皮細胞の3次元像。緑はGFPと融合させた蛋白質、赤はアミロプラストの自家蛍光。 PIN3:オーキシン排出キャリア、mTalin: アクチン繊維結合蛋白質、γ-TIP: 液胞膜蛋白質
これまでの研究から、内皮細胞がシロイヌナズナ花茎の重力感受部位であること、アミロプラストの重力方向への移動が重力感受に重要であること、そして内皮細胞内の液胞やアクチン繊維が花茎の重力感受に関わることを明らかにしました。重力という刺激はオフにすることができないので、生物の重力応答を研究するには特殊な工夫が必要になります。私達は、重力方向を変えるという刺激前後の感受細胞の様子を、生きたままリアルタイムで観察する実験系として、垂直ステージ共焦点顕微鏡や遠心顕微鏡(研究動画)などを独自に立ち上げ、重力の感受メカニズムに迫ろうとしています。
研究動画
4.重力感受を可能にする液胞膜の動的性質
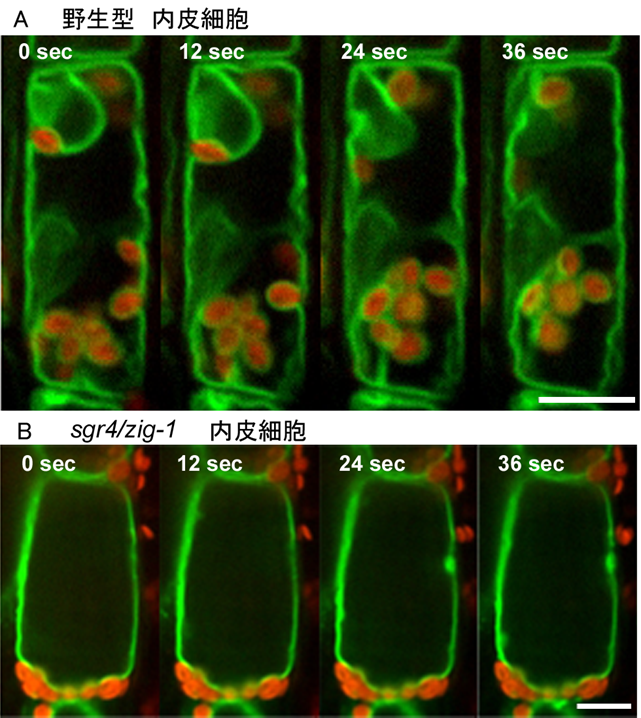
図4 内皮細胞の液胞膜の動的性質とアミロプラスト
共焦点顕微鏡画像の連続写真。A: 野生型、B:zig-1変異体。 野生型では発達した中央液胞が、細胞体積のほとんどを占める。液胞膜の深い貫入(原形質糸)が見られ、アミロプラストの周囲は液胞膜で囲まれたまま、移動している。zig-1変異体では、中央液胞は形成されるが液胞膜の貫入は観察されず、液胞膜がアミロプラストを囲むことは無い。また、アミロプラストもほとんど移動しない。緑:液胞膜、赤:アミロプラストの自家蛍光。
内皮細胞はその体積のほとんどを液胞によって占められ、アミロプラストは周囲を液胞膜に取り囲まれています。私達は変異体の解析を通して、液胞膜が様々に形を変えながら動くという動的性質がアミロプラストの重力方向への移動に重要であることを明らかにしました。液胞は植物個体のあらゆる細胞に存在し多様な役割を担っています。今後、変異体の原因遺伝子の機能を探ることにより、内皮細胞に限らず様々な細胞における液胞膜の動的性質の制御機構、さらには植物が示す重力屈性以外の高次機能において液胞膜動態が果たす未知の役割に迫りたいと考えています。
研究動画
5.重力シグナリングの分子メカニズムに迫る
各器官で重力方向が受容された後、その器官内で成長を調節する植物ホルモンであるオーキシン濃度に差が生じ、器官が屈曲することが示されています。つまり、重力感受細胞においてアミロプラスト沈降という形で受容された重力方向に従って、器官内でのオーキシン分布が制御されることになるわけですが、感受細胞内での情報伝達(重力シグナリング)の仕組みは明らかにされていませんでした(図5)。私たちは、シロイヌナズナを用いて、植物特有の遺伝子LZY (LAZY1-like)ファミリーが、重力感受細胞内で重力刺激に応じたオーキシン偏差分布制御を行うことを明らかにしました。
この研究では、まず重力感受細胞分化不全により花茎の重力屈性能を失った突然変異体eal1と野生型の花茎からRNAを抽出し、マイクロアレイという手法を用いてeal1で発現が低下する遺伝子を探しました。多くの候補遺伝子の中に、イネ地上部の重力屈性異常変異体の原因遺伝子であるLAZY1遺伝子に低い相同性を示す遺伝子が2つ見つかりました。これらをLZY1, LZY2と名付け、LZY2と相同性の高いLZY3と共に、シロイヌナズナにおいて機能解析を行いました。花茎、胚軸、根それぞれの器官で貢献度は異なるものの、LZY1, LZY2, LZY3は重力屈性に冗長的に働くことが分かりました。これらの遺伝子が期待通り重力感受細胞で発現するかを調べたところ、LZY1, LZY2, LZY3は花茎、胚軸において重力感受細胞である内皮細胞で発現し、LZY2, LZY3は根の重力感受細胞であるコルメラ細胞で発現していることが分かりました(図6)。そこで、LZY遺伝子をLZY遺伝子自身以外の重力感受細胞特異的なプロモーター制御下で発現させたところ、LZY遺伝子の多重欠損変異体(lzy多重変異体)の重力屈性異常が回復しました。このことは、LZY遺伝子が重力感受細胞で機能することを強く示しています。重力屈性能を失ったlzy多重変異体では、重力感受細胞は正常に形成され、アミロプラストの重力方向への沈降も正常に起こりますが、重力刺激に応じた器官内でのオーキシン偏差分布が生じないことを見いだしました。従って、LZY遺伝子の機能は、アミロプラスト沈降後、その情報をオーキシン偏差分布へとつなげる感受細胞内での情報伝達であると言えます。
植物は多くの枝や側根を持っており、これらは重力方向に対して一定の角度を保って成長する傾斜重力屈性もしくは横重力屈性と呼ばれる性質を示します。シロイヌナズナの側枝や側根も斜めの角度を保ちますが、lzy 多重変異体では側枝や側根が重力とは逆の方向に斜めの角度を保つという性質を示すことが分かりました(図7)。重力感受細胞特異的にLZY遺伝子を発現させると、正常な角度に戻ることから、主茎や主根の重力屈性と同じ重力情報伝達機構が側枝や側根の角度制御にも使われていることが分かりました。
現在、LZYタンパク質の分子機能の解析を進め、重力シグナリング機構の実体に迫りたいと考えています。
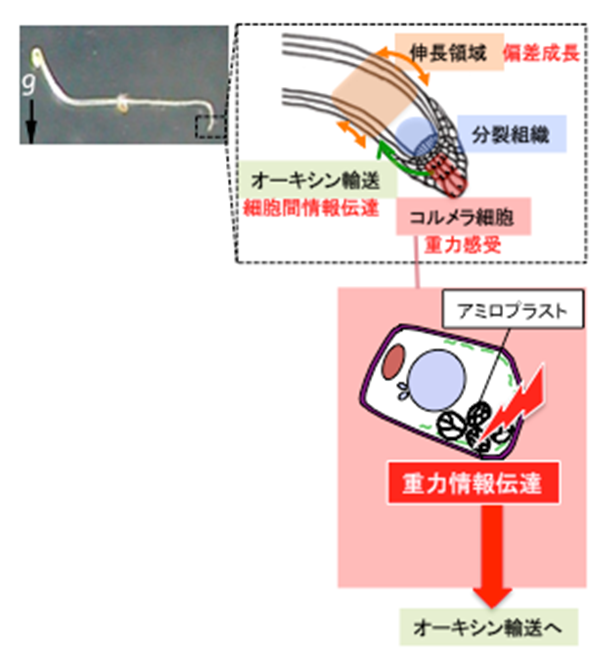
図5 シロイヌナズナの主根における重力屈性反応
重力感受細胞内のアミロプラスト沈降による重力感受から、器官の偏差成長に至る情報の流れを図示した。今回は、感受細胞内での重力情報伝達に関わる遺伝子を見出した。
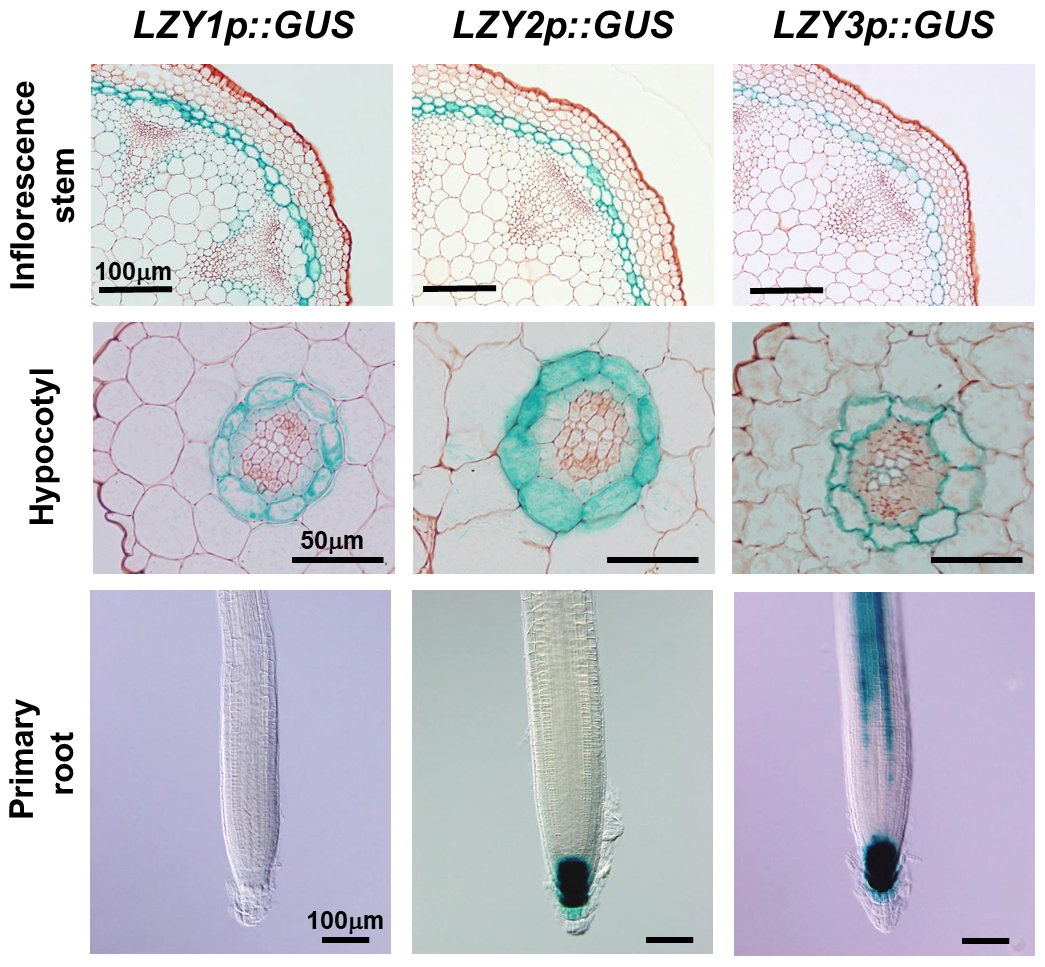
図6 LZY1, LZY2, LZY3 は主に重力感受細胞で発現している
各遺伝子のプロモーター領域(pLZY)にGUS(β-グルクロニダーゼ)遺伝子を連結したものを野生型植物に導入した形質転換体を用いた。各遺伝子が発現する組織が、GUSの働きにより青色に染色される。花茎および胚軸では内皮が、主根では根端のコルメラがよく染色されている。
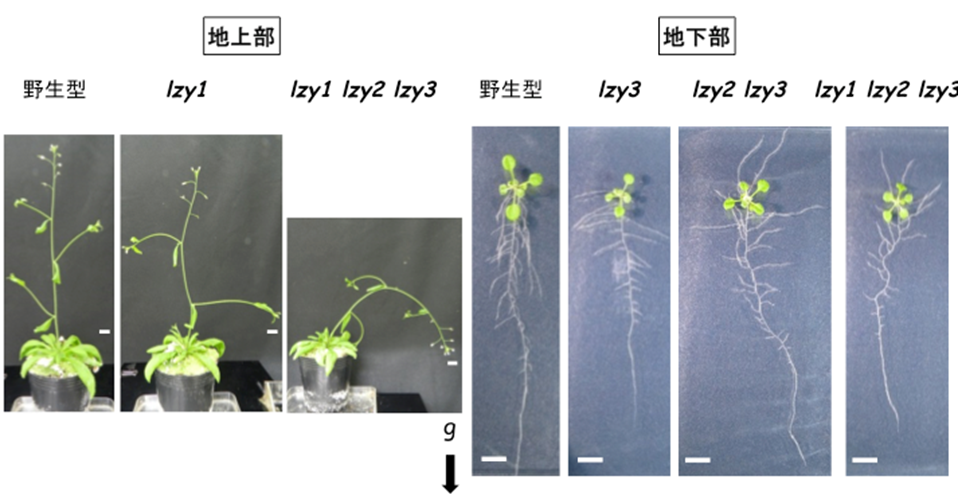
図7 LZYの機能欠損は主軸の重力屈性に加え、側枝及び側根の伸長方向に影響する
lzy1 lzy2 lzy3 三重変異体の側枝は下方に、側根は上方に伸長した。地上部は播種後約6週の植物体、地下部は播種後約2週の植物体。gと矢印は重力の方向を示す。
6.重力屈性とプラントアーキテクチャ

図8
重力屈性と側生器官の伸長角度制御は、植物器官の空間配置によるプラントアーキテクチャ(植物構造:いわゆる枝振り、根の張りや樹冠)をつかさどる要因の1つで、草丈や枝数と同様に育種上重要な形質です。イネやトウモロコシにおいては葉の角度を上に向ける形質が密植や光合成効率の改善につながり、生産量を増大させました。このような農業的重要度に比して、その分子メカニズムの研究は少なく、その理解は進んでいませんでした。しかし、図8に示すように、LZY遺伝子の改変によって、シロイヌナズナの根を重力とは反対の方向に向けることができました。この遺伝子ファミリーの研究をさらに進めることで、将来的には器官の伸長角度を制御してプラントアーキテクチャを改変することができると期待されます。