研究内容(近日大幅更新予定)
脊椎動物の体節形成機構を探る
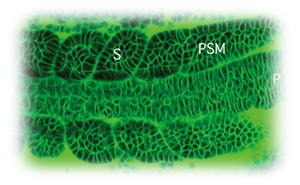
動物のからだには、さまざまな繰り返し構造が認められる。例えば、脊椎は一つ一つの椎骨が連なりあってできている。また、感覚神経節は体幹部において一定の間隔で繰り返し存在する。このような構造の反復性は、もとをたどれば発生初期に一過的に形成される体節の反復性に由来する。脊椎動物の各体節は、発生の進行に従い頭部側から尾部側に向けて順次作られるが、その際、体節は胚の後端に存在する未分節中胚葉から一定の時間間隔のもと、逐次くびれ切れることにより形成される。すなわち、未分節中胚葉において一定の時間間隔のもと繰り返し起きる変化が、体節という形態の反復性を生み出しているわけである。このような「時間的周期性から形態的反復性への変換」は脊椎動物の体節形成を特徴づける大きなポイントとなっており、その変換を生み出す分子メカニズムは興味が持たれるところである
分泌性シグナルタンパク質Wntの分泌・拡散の制御機構を探る
動物の発生過程の様々な局面において、Wnt, BMP, FGFと> いった分泌性のシグナルタンパク質が重要な働きを演じている。これらの分泌タンパク質はその産生細胞自身およびその周囲の細胞に対して働きかけるが、その分泌距離に応じて作用を受ける細胞の数と種類が変わってくることが考えられる。特に、受容する細胞が分泌タンパク質の濃度に応じて作用を異にする場合、すなわち組織の部域化と呼ばれるような現象が引き起こされる場合には、その濃度勾配の傾斜と広がりの程度が組織の形態形成や機能分化に大きな意味をもつ。このような点から考えれば、分泌性シグナルタンパク質が細胞外環境へどのように分泌され、その環境下でどのようにその拡散が制御され、さらにその濃度情報を細胞がどのように認識していくのかという問題は、動物の発生メカニズムを理解する上でまさに不可欠なものであるが、その理解はまだほとんど進んでいない。 Wntタンパク質の分泌にはporcupineと呼ばれる小胞体に局在する 膜タンパク質が必要であることが、ショウジョウバエ、線虫を用いた遺 伝学的解析から示唆されている。 最近porcupineがアシル 基転位酵素と構造的に相同であることが示され、その酵素活性による脂 質修飾がWntタンパク質の分泌に関わることが提唱されてきた。 私たちは独自に確立したWnt3aタンパク質を効率良く細胞外に分 泌する培養細胞系(Shibamoto et al. 1998)を用いて、 Wnt-3aタンパク質が不飽和脂肪酸による独特の脂質修飾を受けること、 そしてこの修飾にはporcupineが必要であることを示すととも に、この修飾はWnt-3aタンパク質が小胞体から輸送される上で必 要であることを明らかにした(Takada et al. in press)。現在 は、この結果をもとにして、Wntタンパク質の分泌、拡散の機構 の解明に向けたさらなる研究を続けている。
シグナルタンパク質の機能多様性が生み出される仕組みを探る
近年の発生生物学研究により明らかにされた重要な知見の一つは、細胞外に分泌されるシグナルタンパク質が動物の形態形成過程において実に多種多様なはたらきをするということである。Wnt、FGF、BMPそれにHedgehogといったシグナルタンパク質が、細胞の増殖、分化、移動、さらには組織の領域形成など形態形成に関わるさまざまな現象に関与することがこれまでに示されてきた。そこから浮かび上がってくる個体の形態形成メカニズムの全体像は、限られた種類のシグナルが時期と場所を変えてくり返し使われ、しかもその作用は各々のケースごとに異なり、極めて多彩であるというものである。 このような全体像を前にした時、一つの問題が生じる。すなわち、「くり返し使われる同じシグナル(単一のインプット)からどのようにして受け手の細胞(以下、標的細胞と呼ぶ)ごとに異なる作用(複数のアウトプット)が生じるのであろうか?」ということである。この問題は個体の形態形成メカニズムを理解していく上で根源的なものであると思われるが、今までのところ標的細胞が内在的に持つシグナルに対する応答性の違い(または、「コンテクストの違い」)といった観念的な言葉でのみ説明されているにすぎず、分子レベルでの解析は進んでいないのが現状である。 私たちは、このようなシグナルの機能の多様性が生じる分子メカニズムを解析することを目的として、シグナルによって発現が調節される遺伝子(以下、標的遺伝子と呼ぶ)の発現制御機構に着目している。マウスの初期発生過程においてWntシグナルは原口(原始原条)周囲や神経管背側といった複数の領域で活性化されており、その機能もまた多岐にわたる(Takada et al., 1994, Yoshikawa et al.,1997, Ikeya et al., 1997; Ikeya & Takada, 1998; Muroyama et al. 2002)。このうち、原口周囲においてはWntシグナルによって転写調節因子であるT/Brachyuryが転写活性化されることがすでに示されている(Yamaguchi et al., 1999)。しかしながら、T/BrachyuryはWntシグナルが活性化されているその他の領域では発現しておらず、この遺伝子の発現がWntシグナルに対する原口周囲特異的な応答性をモニタリングする上での良い指標となる。 私たちは、ゼブラフィッシュを用いてT/Brachyuryを異所的に発現する変異体を多数獲得することに成功した。興味深いことにこれら変異体においてはT/Brachyuryが本来の発現領域である原口周囲等の他に、予定中脳・後脳境界領域で発現している。予定中脳・後脳境界領域では、原口周囲と同様にWntやFGFといったシグナル分子が発現することから、このような変異体ではこれらのシグナルによるT/Brachyuryの発現誘導が原口周囲以外の領域でも起きるようになったのではないかと考えられる。現在、これら変異体の原因遺伝子を同定し、それらがコードする遺伝子産物の分子機能を明らかにすることにより、シグナルに対する応答性の違いを生み出すメカニズムの解明に迫りたいと考えている。 また、これと平行して、Wntシグナルの標的遺伝子の探索を行っている。これまでのところ、マウス胚性幹細胞(ES細胞)に対してWnt-3aタンパク質を添加することによって発現が変動する遺伝子として、10数個の標的遺伝子の同定に成功した。さらに、そのうちのいくつかはマウスの発生過程においてWntシグナルが機能することが明らかになっている細胞・組織で発現することも示されている(Yamaguchi et al., 2005)。現在、遺伝子トラップしたマウスES細胞もしくは新たに遺伝子ターゲッティングしたES細胞から標的遺伝子の変異体個体を作製し、発生過程においてWntシグナルが担う多様な機能のうちのいったいどこにこれら標的遺伝子が関与するのかを明らかにしようとしている。