領域代表
北沢 美帆
大阪大学 全学教育推進機構 講師
地球上には多様な形態を示す生物が進化してきました。なかでも、植物や大型藻類など、自発的な移動能力を持たない固着性多細胞生物には顕著な形態的多様性がみられ、たとえ遺伝子レベルでは同一であっても全く異なった形態をつくりだします。例えば、タンポポの葉は、日当たりなどの生育環境によって、切れ込みの数や間隔・深さに可塑性を示します。キンポウゲ科の花弁や萼片の数は、同一環境の個体群、個体内でも異なることがあり、遺伝子発現の確率性によると考えられています(Kitazawa & Fujimoto, 2014; Wang et al., 2015)。
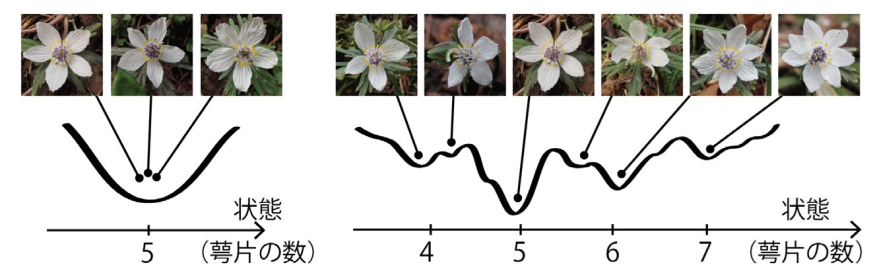
近年、このような「同一の遺伝型から生じる表現型のゆらぎ」と形質進化が正に相関することが提案されています(Kaneko & Furusawa 2006)。しかし、多くの実験系では、ゆらぎは「個体差」や「ノイズ」として、平均化され、表現型の議論から排除されてきました。自然界にみられる多彩なゆらぎの理解は、「それぞれの形質は特定の遺伝子によって決まる」「可塑性は、少数の状態の間を特定の遺伝子の発現によってスイッチすることで生じる」といった従来の視点のみでは困難です。技術の発展により非モデル生物を対象とする研究や野外環境での様々な研究が可能となり、生物多様性の理解に対する社会的要請も高まっている今、形態の柔軟性に対する新たな視点からの研究が必要とされています。
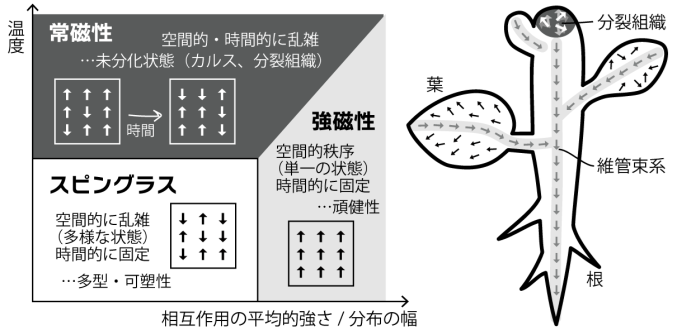
本領域は、陸上植物や大型藻類など固着性の多細胞生物の形態の柔軟性が、細胞レベルでの非一様性によって創発することを提案します。この理論的背景は、ミクロな非一様性がマクロな特性を創発するスピングラス理論です。
一般に、金属など磁性体の全体的な特性は、ミクロなスピンの向き・並び方に左右されます。すべてのスピンが同じ向きにそろう単純な場合は、全体の極性も単純に定まる(強磁性)。一方、ミクロなスピンがバラバラな向きのまま固定されたスピングラスは、多数の安定状態など強磁性体には見られない特性を示します。ここから、ミクロな要素の非一様性は、全体としての新たな特性を創発することが示唆されます(西森秀稔, スピングラスのゲージ理論, 1988)。
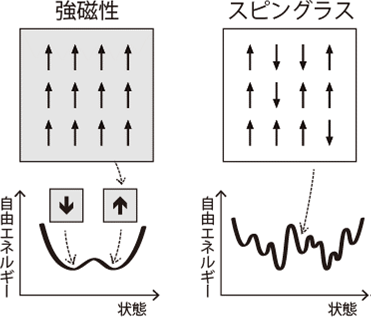
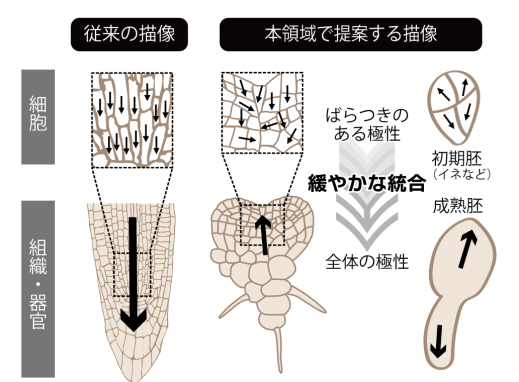
植物の発生過程には、ミクロな極性のばらつきが維持されながらも、発生の進行にともない安定状態を形成する「スピングラス」的状態がみられます。その具体例として、植物の発生において中心的役割を果たすオーキシンの分布が挙げられます。オーキシンの分布を主として決定する輸送担体PINの細胞における偏在は、器官新生の初期は緩やかであり、発生の進行にともない自己組織的に整っていきます。スピングラス理論に従えば、個々の細胞の性質や細胞間相互作用がばらつき(分布)を持つとき、全体としての新たな性質が生まれます。我々はこの概念を基に,従来は軽視・排除されていた「個体差」や「ノイズ」を含め発生様式を統合的に理解する新たな学術領域として『植物スピングラス』領域を創成しました。