種子貯蔵タンパク質の蓄積機構の解析
成長した植物は、次世代を残すために種子を形成します。種子は、多量の脂質やタンパク質、糖質を蓄積し、種子重量の大半がそれら種子貯蔵物質で占められます。脂質は小胞体由来のオルガネラであるオイルボディに、タンパク質は液胞由来のオルガネラであるプロテインボディに蓄積し、種子細胞のほとんどがオイルボディとプロテインボディで占められます (図1)。この蓄えられた貯蔵物質の品質や量、タンパク質と脂質の割合が、食糧や材料としての種子の品質を決定しています。このため、種子における貯蔵物質の合成と蓄積機構の解明は、より質の高い種子の生産につながります。
|
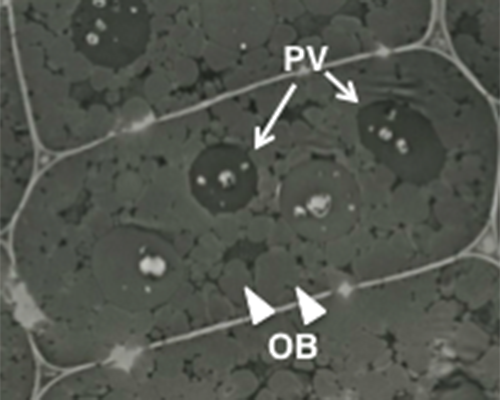
| 図 1. シロイヌナズナ種子の電子顕微鏡写真。
タンパク質を蓄積しているプロテインボディ (PV、矢印) と脂肪を貯めているオイルボディ (OB、矢頭) で占められている。 |
私達は、種子における脂質合成の制御機構の解明を進めています。脂肪性種子植物であるシロイヌナズナを用いて、種子の密度を指標に種子中の脂質含量が変化している変異体を選抜し、原因遺伝子の同定とコードするタンパク質の機能解析を行っています。これまでに、種子の密度が高くなっている変異体 (野生株よりも脂質蓄積量が低下している変異体) として単離されたhs3(heavy seed 3)の解析から (図2)、HS3遺伝子が葉緑体に局在するDNA/RNAヘリカーゼをコードし、ACCaseなどの葉緑体ゲノムの遺伝子発現に関与していることを明らかにしています。
|
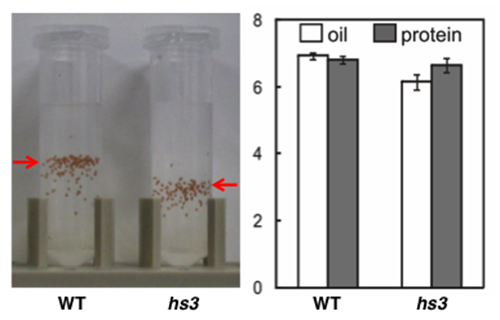
| 図 2. heavy seed 3 (hs3) 変異体では、脂質とタンパク質の蓄積が変わっている。
野生型 (WT) とhs3変異体の種子を、パーコール溶液で遠心分離すると、hs3変異体では、WTよりも下にきます。種子1粒あたりの脂質とタンパク質含量を定量すると、hs3変異体ではタンパク質含量はWTと変わらないものの、脂質含量が、10〜15%程度減少していることが明らかとなった。 |
植物では、葉緑体で合成された脂肪酸が小胞体に輸送され、オイルボディの構成成分であるトリアシルグリセロールが作られます。小胞体に蓄積したトリアシルグリセロールは、オイルボディとして切り離されます。このように脂質合成には、葉緑体、小胞体、オイルボディという3つのオルガネラが関与しており、HS3は植物の初期生長、および種子の脂質貯蔵合成に関与していることが分かってきました (図3)。
hs変異体とは逆に種子密度が低くなっている変異体も得られていますので、これらと他のhs変異体の解析を進め、種子貯蔵タンパク質の蓄積機構の解明に取り組んでいます。
|
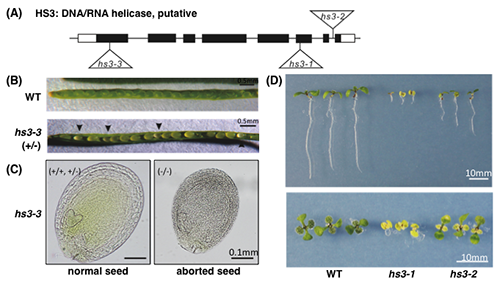
| 図 3.hs3変異体の解析
HS3遺伝子は、葉緑体に局在するDNA/RNAヘリカーゼをコードしている。hs3変異体には3つのT-DNAが挿入された変異体がある (A)。第1エクソンにT-DNAが挿入されたhs3-3ではホモ個体が得られず、ヘテロ接合体を自家受粉させたさやでは25%の割合で異常な種子が観察される (B)。正常な種子では、正常な胚発生が観察されるが (C, normal seed)、異常な種子では球状胚のステージで停止しており、緑色も観察されないことから (C, aborted seed) 、hs3-3は胚発生致死となることが分かる。一方、hs3-2では、野生株よりは生育が遅れるものの、hs3-1に比べ生育遅延は弱いことが明らかとなった。これらの結果は、HS3が胚発生、及び発芽後の初期生長に必須であることを示している。 |