《 基礎生物学研究所要覧 》統合バイオサイエンスセンター
|
| DEPARTMENT OF DEVELOPMENT, DIFFERENTIATION AND REGENERATION II |
|
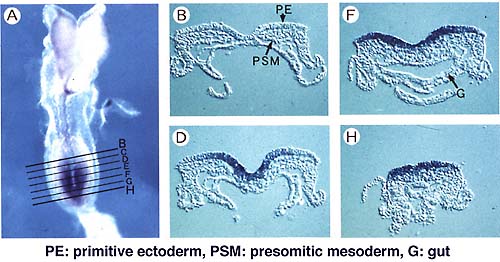
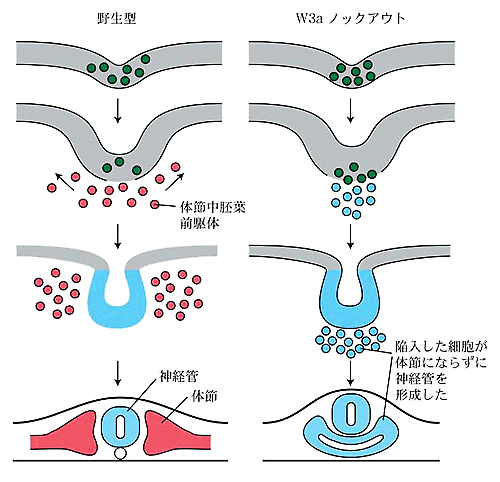
1.脊椎動物の体節形成におけるWntシグナルのはたらき近年、WntやFGFをはじめとする分泌性タンパク質が動物の形態形成過程において重要な働きを担うことが示されてきた。Wnt遺伝子は線虫から脊椎動物にいたるはば広い動物種に存在し,発生過程においてさまざまな役割を演じている。また,各々の動物種においては多数のWnt遺伝子が存在することも報告されており,脊椎動物では少なくとも17個のWnt遺伝子の存在が確認されている。 そのうちの一つWnt-3aは,マウスの初期発生過程では体幹部の後部末端に存在する原条もしくは尾芽と呼ばれる領域に発現する(図1a)。この領域の細胞は体節を含む体幹部の形態形成に中心的な役割をになうものと考えられているが、興味深いことに,Wnt-3a遺伝子の機能欠失型変異体マウス胚では体節前駆細胞が神経管に分化転換してしまう(図1b)。このことから,Wnt-3aシグナルは体節前駆細胞の分化経路の正しい選択に必須であること,また,体節前駆細胞は神経上皮細胞に分化できる潜在能力を持っているもののWnt-3aシグナルによりその分化方向が規定されているものと考えられた。一方,上記の異常とは別に,Wnt-3a変異体においては各体節の特徴が部分的に前方化もしくは後方化していることも見い出されており,Wnt-3aは前後軸に沿った各体節の特徴づけにも関与しているものと考えられる。このようなことから,Wnt-3aは体節の発生過程において複数の役割をはたしていると思われる。
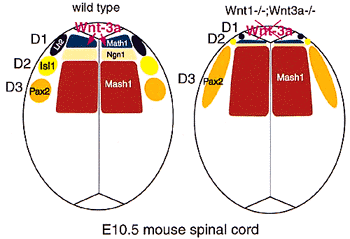
そこで,Wntシグナルにより制御される体節形成の各々の過程の分子機構を理解するために,現在,Wntシグナルによりその発現が制御される遺伝子の探索を始めており,そのような遺伝子の機能解析を通して体節前駆細胞におけるWntシグナルの作用機構を明らかにしていきたいと考えている。また,体節形成過程におけるWntシグナルに対する応答能の制御機構を調べる目的で,体節前駆細胞で発現するWnt受容体を同定しており,この受容体とWntとの相互作用等についても解析を進めている。一方,脊椎動物における体節形成の分子機構を明らかにする別のアプローチとして,遺伝学的な解析が可能なゼブラフィッシュを用いて体節形成に異常を呈する変異体のスクリーニングも試みている。さらに,それと平行して,ゼブラフィッシュ胚の尾芽で発現する遺伝子の網羅的スクリーニングも行っており,尾芽における体節中胚葉の発生機構を多面的なアプローチにより解明しようとしている。 2.神経管背側におけるWntシグナルのはたらきWnt-3aはWnt-1とともに胎生期のマウスの神経管の背側領域領域においても発現する(図2a)。Wnt-1とWnt-3aの二重変異体においてはこの領域から発生する神経冠細胞の数に減少が認められる(図2b)。また、神経管の両側に位置する体節の発生にも神経管背側から分泌されるWnt分子は重要な働きをしており,体節の中で筋分化に重要な働きをする皮筋節の内側部(medial lip)の形成に神経管の背側領域からのWntシグナルが必須であることが示されている。このような機能の他に,神経管背側から分泌されるWntシグナルは,背側介在神経の領域特異化を制御していることを最近明らかにした。現在、神経分化に及ぼすWntシグナルの効果についても解析を進めている。 3.骨形成におけるFGFシグナルのはたらきマウスの体節形成過程ではいくつかのFGF遺伝子が発現するが,そのうちの一つであるFGF18遺伝子の変異体を作成したところ,体節の発生は正常であり、むしろ骨形成に異常が認められた。詳細な解析を行った結果,FGF18には軟骨形成は負に,骨形成は正に制御するという二面的な機能があることがわかった。脊椎動物の発生が秩序だって進行するためには骨が他の組織と調和を保って形成されていくことが重要であり,そこでは骨細胞の分化と増殖は厳密に制御されているものと考えられる。そこで,現在,FGF18による骨形成の分子機構の解析を行っている。
参考文献
|
| Copyright(C) National Institute for Basic Biology (webmaster@nibb.ac.jp) | ||