《 基礎生物学研究所要覧 》形質統御実験施設
|
| DIVISION OF SPECIATION MECHANISMS II |
|
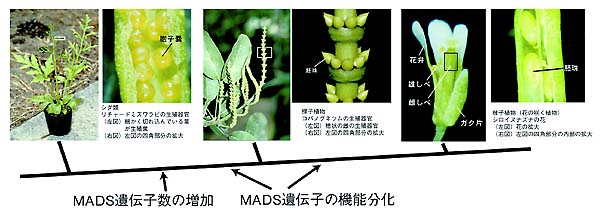
現存する全ての生物は約40億年前に生じた1つの共通祖先から進化してきた。従って,現生生物に見られる多様性は,40億年間に蓄積した突然変異によって引き起こされたものである。そして,生物の進化過程の痕跡は,現生生物のゲノム上に記されている。異なった生物間で,ゲノムの配列情報,および,ゲノム情報によってうみ出される遺伝子の働きを比較解析することにより,どのように進化がおきてきたかを解明することができる。 我々は,まず,(1)生物の正しい類縁,系統関係を遺伝子配列から推定し,(2)得られた系統樹からどのような傾向で形態形質が進化したかを解明し,さらに(3)生物の形態の進化がどのようなゲノム上の変化によって引き起こされたのかを明らかにしようとしている。 1.花の進化を探る花は植物の生殖器官である。花はがく片,花弁,雄しべ,雌しべの4つの花器官からできており,雄しべと雌しべの中で減数分裂により生殖細胞が形成される。一方,より原始的なシダ類では,生殖細胞は胞子嚢と呼ばれる1重の袋に覆われ,葉の裏にむきだしについており,より単純な形をしている。では,どのような変化がおこって,シダ類のような単純な生殖器官から花が進化してきたのだろうか。 花の形態形成に関係する遺伝子が花の咲く植物で解析され,MADS-box遺伝子群と呼ばれる転写調節因子が花器官形成に深く関与していることが明らかになってきた。では,花の咲かないシダ類にはこの遺伝子群は存在しているのであろうか,それともこの遺伝子の誕生が花の進化に関わったのであろうか。我々は,シダ類の中で世代時間が短く新しいモデル植物として着目されているリチャードミズワラビからMADS-box遺伝子を単離することに成功した。その結果,リチャードミズワラビもMADS-box遺伝子を持っていることがわかった。しかし,花の咲く植物では,10以上ものMADS-box遺伝子のグループがあるのに,リチャードミズワラビには3つ程度のMADS-box遺伝子グループしか存在していないらしいことがわかった。さらに,花の咲く植物では,それぞれのMADS-box遺伝子は特定の器官でのみ発現し,特定の器官形成に関わっていることが多いのに対し,シダ類のMADS-box遺伝子の発現は,特定の器官ではなく,生殖器官,栄養器官の両方で広範に発現しており,MADS-box遺伝子の機能が未分化であるらしいこともわかった。このことから,シダ類のような原始的植物で,生殖,栄養両器官の形態形成にかかわっていたMADS-box遺伝子の(1)数が増え,(2)増えて余った遺伝子がそれまで発現していなかった特定の場所で発現するようになり,花器官を進化させた,というシナリオが描けた。現在,コケや緑藻類など,より原始的な植物におけるMADS-box遺伝子の機能解析から,花器官形成遺伝子の進化の全貌を明らかにすることを目指している。
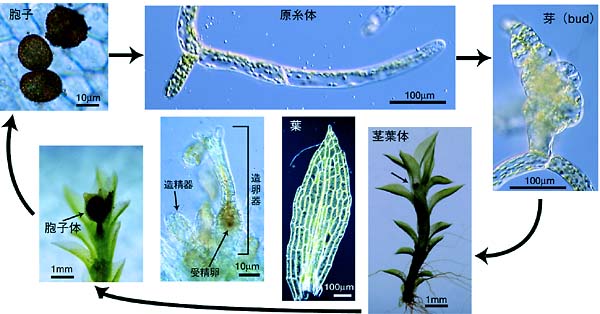
2.植物の分裂組織形成,維持,器官形成メカニズムと進化屋久島の屋久杉は何千年も生き続けている。これは,植物の体が茎頂の分裂組織からたえず作られ続けており,その分裂組織が半永久的に成長し続けるからである。 植物の地上部のほとんどは茎の先端にある茎頂分裂組織から形成される。被子植物では数細胞層からなる多細胞性の茎頂から順次,規則正しく,多細胞性の葉と茎が形成されてくる。一方,コケ植物の蘚類は,2つの異なった細胞分裂機構を持っている。茎葉をつける茎葉体では,単細胞の茎頂分裂細胞から,それぞれ1細胞性の葉と茎の原基細胞が形成され,それぞれが多細胞性の葉と茎へと分化していく。さらに,糸状の体制を持った原糸体では,頂端分裂細胞が2分裂することにより,1次元的に細胞の糸が作られて行く。前者は被子植物など高等植物の持つ分裂様式このように異なった分裂組織は,どのように進化してきたのだろうか。 植物の分裂組織に関する研究は,花器官形成ほど進展していないので,まず,茎頂分裂組織形成,維持,器官形成の分子機構自体から解明していく必要がある。そのためのモデルとして,我々はコケ植物蘚類のヒメツリガネゴケを選んだ。ヒメツリガネゴケは,陸上植物では唯一,高い相同組換え率を持っており,遺伝子ターゲティングが容易である。まず,ヒメツリガネゴケ遺伝子トラップ系を用いて茎頂分裂組織特異的に発現している遺伝子を探し,その機能解析をすすめている。さらに,原糸体、茎葉体由来のRNAから,全長cDNAライブラリーを作成し,そのEST解析を行うことにより,約1万5千の独立した,全長cDNAをカタログ化した。これらのcDNAをヒメツリガネゴケで網羅的に過剰発現させることにより分裂組織形成・維持に関わる遺伝子の単離を目指している。 参考文献
|
| Copyright(C) National Institute for Basic Biology (webmaster@nibb.ac.jp) | ||