《 基礎生物学研究所要覧 》制御機構研究系
|
| DIVISION OF MOLECULAR NEUROBIOLOGY |
|
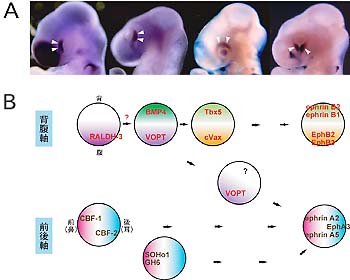
1.網膜における領域特異化の分子機構脳・神経系では,領野,神経核等と呼ばれる数多くの領域区分が存在し,それぞれ独自の機能を担っている。しかしながら,その形成の仕組みは未だ充分に解明されていない。我々は,脳の一部から発生する眼における領域特異化の問題を取り上げ,網膜において前後軸(鼻耳軸)並びに背腹軸方向の領域特異性獲得の分子機構を明らかにする研究を行っている。既にRLCS法によって,ニワトリ胚の網膜において領域特異的に発現する分子群を網羅的に単離・同定する作業を完了した(図1A)。同定した分子は前後軸方向で33分子,背腹軸方向で20分子におよぶが,この中には数多くの転写調節因子,膜分子,分泌因子,シグナル伝達因子,細胞骨格関連分子等が存在している。これまでに、CBF-1,-2等の転写調節因子,RALDH-1,-3等のレチノイン酸合成酵素,Ventroptinと名付けたBMP中和因子等を同定し報告している。引続き,異所的な遺伝子発現,遺伝子変換マウスの作製等によって,これらの遺伝子の機能と相互関係を解析する(図1B)。この研究を通して,眼のできる仕組み,網膜における領域特異化の仕組みを解明する。
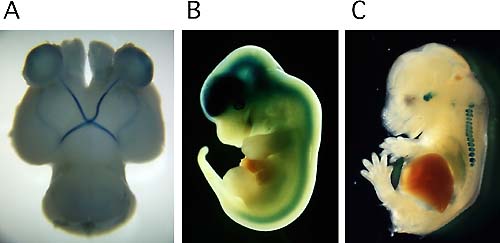
2.領域特異的神経結合形成の分子機構神経系では,その発生過程において様々な領域で,ある部域の神経細胞から発した神経軸索が,別の特定の領域の神経細胞に対して,二次元的相対位置関係を保存した形で正確に対応して結合する,いわゆるトポグラフィック投射路が形成される。ニワトリの網膜視蓋投射の系では,網膜の鼻側(前側)あるいは耳側(後側)の領域から発した視神経は,視中枢(視蓋)のそれぞれ後側,前側の領域に選択的に神経結合を形成する。同様に,背側から腹側,腹側から背側の領域に投射が起こる。この視神経のトポグラフィックな投射には,上記の網膜の領域特異化が密接に関係している。我々は,網膜において領域特異的な発現を示す転写調節因子等の発現部位を変えることによって,視神経の投射先部位を人意的に変えうることを示した。ニワトリとマウスを用いて,視神経が正しい相手と神経結合を形成する仕組み,特に軸索先端の成長円錐の挙動をコントロールする分子機構を中心に研究を行っている(図2A)。 3.プロテオグリカン型チロシンホスファターゼPTPζの役割中枢神経系における主要な細胞外基質分子はプロテオグリカンであり,いくつかの分化(栄養)因子は,プロテオグリカンのグリコサミノグリカン鎖に結合することによって,初めて機能的なリガンドとなることが示されている。 我々は,受容体型チロシンホスファターゼ(RPTP)の中にプロテオグリカンに属する分子が存在すること,またその内の一つがPTPζ(RPTPβ)であることを明らかにした。また,PTPζには3つのスプライシングアイソフォームが存在することを示した。更に,このPTPζのリガンド分子としてPleiotrophinとMidkineを同定し,これらリガンドの結合により神経細胞分化,細胞移動が誘導されることを見い出した。また最近,PTPζが,C末でPSD95ファミリーと結合していること,またGIT1を基質とすることを見い出し,シナプス伝達の調節にも関与している可能性が明らかになってきた。今後,GIT1以外の細胞内基質分子の同定を進めると共に,PTPζ遺伝子ノックアウトマウスの解析によって,本分子の情報伝達経路の究明と脳形成,脳機能における役割に迫る(図2B)。
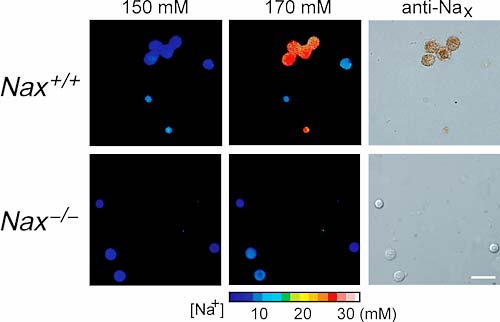
4.Naxチャンネルの機能これまでNaG(SCL11),Nav2.1, Nav2.3等と呼ばれてきたNaxイオンチャンネルは,電位依存性Naチャンネルファミリー(Nav1)と一次構造上,比較的近い構造を有するものの,その機能と役割は明らかになっていなかった。我々は,この遺伝子欠損マウスを作製し(図2C),その解析を通して,このチャンネルが体液中の塩濃度検知に関わる脳室周囲器官に発現していること,欠損マウスでは脳室周囲器官の活動が活性化されていること,また,食塩水を異常に摂取することを見い出した。更に最近,このチャンネルが,細胞外のNa+イオン濃度の生理的範囲での上昇(150→170mM)を検知して開くNaチャンネルであることを明らかにした(図3)。今後は体液塩濃度の恒常性維持に関わる脳内機構を明らかにする研究を展開する。
図3.脳弓下器官ニューロンのNa+流入応答 参考文献
|
| Copyright(C) National Institute for Basic Biology (webmaster@nibb.ac.jp) | ||