| DIVISION OF MOLECULAR NEUROBIOLOGY | ||||||||
|
||||||||
| 1) CREST, JST 2) PREST, JST | ||||||||
We have been studying the molecular
and cellular mechanisms underlying the development of the vertebrate
central nervous system. It covers all the major events including the
patterning of the nervous system, neuronal differentiation, axonal
navigation and targeting, synapse formation and plasticity, and neuronal
regeneration, mainly in the visual system. The scope of our interests
also encompasses various functions of the matured brain, including
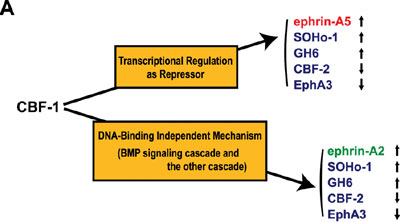
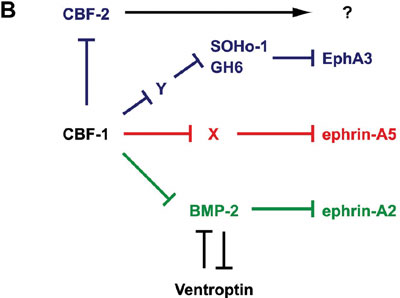
sensation, behavior, learning and memory. Topographic maps are a fundamental feature of neural networks in the nervous system. Understanding the molecular mechanisms by which topographically ordered neuronal connections are established during development has long been a major challenge in developmental neurobiology. The retinotectal projection of lower vertebrates including birds has been used as a readily accessible model system. In this projection, the temporal (posterior) retina is connected to the rostral (anterior) part of the contralateral optic tectum, the nasal (anterior) retina to the caudal (posterior) tectum, and likewise the dorsal and ventral retina to the ventral and dorsal tectum, respectively. Thus, images received by the retina are precisely projected onto the tectum in a reversed manner. Since 1992, we have been devoting our efforts to searching for molecules with asymmetrical distribution in the embryonic chick retina, and to characterization of their roles in the topographic retinotectal projection. In 1996, we first identified two winged-helix transcriptional regulators, CBF-1 and CBF-2, expressed in the nasal and temporal retina, respectively. Misexpression experiments in the retina using a retroviral vector showed that these two transcription factors determine the regional specificity of the retinal ganglion cells, namely, the directed axonal projections to the appropriate tectal targets along the anteroposterior axis. Secondly, we identified a novel retinoic acid-generating enzyme, RALDH-3, which is specifically expressed in the ventral region of the retina, together with a dorsal-specific enzyme RALDH-1. Furthermore, we recently identified a novel secretory protein, Ventroptin, which has BMP-4 neutralizing activity. Ventroptin is expressed in the retina with a ventral high-dorsal low gradient at early stages. This expression pattern is complementary to that of BMP-4. At later stage (E6~), a nasal high-temporal low gradient expression pattern of it is also detected. Ventroptin thus shows a double-gradient expression profile along the dorsoventral and nasotemporal axes. Misexpression of Ventroptin altered expression patterns of several topographic genes and projection of the retinal ganglion-cell axons to the tectum along the both axes. In this year of 2003, we revealed that misexpression of CBF-1 represses the expression of EphA3 and CBF-2, and
induces that of SOHo-1, GH6, ephrin-A2 and ephrin-A5. CBF-1 controls ephrin-A5 by a DNA binding-dependent mechanism, ephrin-A2 by a DNA
binding-independent mechanism, and CBF-2, SOHo-1, GH6 and EphA3 by dual mechanisms (Fig. 1A). BMP-2 expression begins double-gradiently in the retina from E5 instead
of BMP-4 in a complementary pattern to the Ventroptin expression. Ventroptin antagonizes BMP-2 as well as BMP-4. CBF-1 interferes
in BMP-2 signaling and thereby induces expression of ephrin-A2.
Our data suggest that CBF-1 is located at the top of the
gene cascade for the regional specification along the nasotemporal
(NT) axis in the retina (Fig. 1B) and distinct BMP signals play pivotal
roles in the topographic projection along both axes.
Currently, with respect to the other identified molecules, we are
conducting misexpression experiments using chick embryos and generating
knockout and transgenic mice to elucidate the molecular functions.
We expect that our studies will lead to dissection of the molecular
mechanism underlying the retinal patterning and topographic retinotectal
projection, and ultimately to uncovering the basic principles for
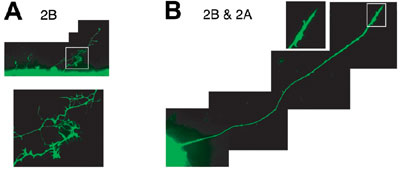
establishing complicated but extremely precise neural networks. During development, cells undergo dynamic morphological changes by rearrangements of the cytoskeleton including microtubules. However, molecular mechanisms underlying the microtubule remodeling between orientated and disorientated formations are almost unknown. We found that novel subtypes of collapsin response mediator proteins (CRMP-As) in addition to the originals (CRMP-Bs), which occur from the alternative usage of different first coding exons, are involved in this conversion of microtubule patterns. Overexpression of CRMP2A and CRMP2B in chick embryonic
fibroblasts induced orientated and disorientated patterns of microtubules,
respectively. Moreover, sequential overexpression of another subtype
overcame the effect of the former expression of the countersubtype.
Overexpression experiments in cultured chick retinae showed that CRMP2B
promoted axon branching and suppressed axon elongation of ganglion
cells, while CRMP2A blocked these effects when co-overexpressed (Fig.
2). Our findings suggest that the opposing activities of CRMP2A and
CRMP2B contribute to the cellular morphogenesis including neuronal
axonogenesis through remodeling of microtubule organization.
Protein tyrosine phosphorylation plays crucial roles in various biological aspects including all stages of brain development. In 1994, we found that PTPz/RPTPb/Ptprz, a nervous system-rich receptor-type PTP, is expressed as a chondroitin sulfate proteoglycan in the brain. Ptprz is expressed from the early developmental stage to the adulthood in neurons as well as astrocytes. This suggests that this molecule plays variegated roles in the brain development and brain function. We found in 1996 that Ptprz binds pleiotrophin (PTN)/HB-GAM and midkine (MK), closely related heparin-binding growth factors which share many biological activities. The chondroitin sulfate portion of Ptprz is essential for the high affinity binding (Kd = ~0.25 nM) to these growth factors, and removal of chondroitin sulfate chains results in a marked decrease of binding affinity (Kd = ~13 nM). To identify the substrate molecules of Ptprz, we recently developed the yeast substrate-trapping system. This system is based on the yeast two-hybrid system with two essential modifications: Conditional expression of v-src to tyrosine-phosphorylate the prey proteins and screening using a substrate-trap mutant of PTP as bait. Using this system, we successfully isolated a number of candidate clones for substrate molecules (ex. GIT1/Cat-1) and continuously-interacting molecules (ex. PSD-95/SAP90) for Ptprz. We are now continuing efforts to characterize these candidate clones. In addition, to know the physiological roles of Ptprz in vivo,
we generated Ptprz-deficient mice in which the Ptprz gene
was replaced with the LacZ gene in 1997. We are currently
studying the phenotype of Ptprz-deficient mice biochemically,
anatomically, physiologically and ethologically.
Sodium-level sensing system in the brain is essential for the regulation of the sodium and water balance in body fluids. Previously, we demonstrated that Nax (also called Nav2/NaG) gene is expressed in the circumventricular organs and Nax-deficient mice ingest salt in excess. In Nax-deficient mice, c-fos expression in the subfornical organ (SFO) and organum vasculosum laminae terminalis (OVLT) was markedly elevated as compared with wild-type mice. Stimulation of the SFO/OVLT of wild-type animals by infusion of a hypertonic sodium solution into intracerebroventricule (ICV) leads to avoidance of salt intake. In contrast, Nax-deficient mice did not show such aversion. We showed that Nax is a sodium channel which is sensitive
to the increase of extracellular sodium level. Entry of sodium ions
occurred in response to a rise of the extracellular sodium concentration
(C1/2= 159 mM). In contrast, these responses
were not observed in Nax-immunonegative cells or neurons
of Nax-deficient mutant origin. Transfection
of Nax cDNA conferred the sodium sensitivity
on Nax-deficient cells. All of the GABA-immunopositive
neurons isolated from the SFO responded to the extracellular sodium
increase. Based on these findings, we proposed that GABAergic inhibitory
neurons expressing Nax control the activity
of the SFO and suppress the salt-intake behavior of animals under
thirst conditions. |
||||||||
Publication List: Asahi, M., Tanaka, Y., Izumi, T., Ito, Y., Naiki, H., Kersulyte, D., Tsujikawa, K., Saito, M., Sada, K., Yanagai, S., Fujikawa, A., Noda, M. and Itokawa, Y. (2003) Helicobacter pylori CagA containing ITAM-like sequences localized to lipid rafts negatively regulates VacA-induced signaling in vivo. Helicobacter, 8, 1-14. Fujikawa, A., Shirasaka, D., Yamamoto, S., Ota, H., Yahiro, K., Fukada, M., Shintani, T., Wada, A., Aoyama, N., Hirayama, T., Fukamachi, H. and Noda, M. (2003) Mice deficient in protein tyrosine phosphatase receptor type Z are resistant to gastric ulcer induction by VacA of Helicobacter pylori. Nature Genet., 33, 375-381. Nakayama, M., Kimura, M., Wada, A., Yahiro, K., Ogushi, K., Niidome, T., Fujikawa, A., Shirasaka, D., Aoyama, N., Kurazono, H., Noda, M. and Hirayama, T. (2003) Helicobacter pylori VacA activates the p38/ATF-2-mediated signal pathway in AZ-521 cells. J. Biol. Chem., (Online in Nov.) Sakaguchi, N., Muramatsu, H., Ichihara-Tanaka, K., Maeda, N., Noda, M., Yamamoto, T., Michikawa, M., Ikematsu, S., Sakuma, S. and Muramatsu, T. (2003) Receptor-type protein tyrosine phosphatase z as a component of the signaling receptor complex for midkine-dependent survival of embryonic neurons. Neurosci. Res., 45, 219-224. Tanaka, M., Maeda, N., Noda, M. and Marunouchi, T. (2003) A chondroitin sulfate proteoglycan PTPz/RPTPb regulates the morphogenesis of Purkinje cell dendrites in the developing cerebellum. J. Neurosci., 23, 2804-2814. Watanabe, U., Shimura, T., Sako, N., Kitagawa, J., Shingai, T., Watanabe, E., Noda, M. and Yamamoto, T. (2003) A comparison of voluntary salt-intake behavior in Nax-gene deficient and wild-type mice with reference to peripheral taste inputs. Brain Res., 967, 247-256. Yuasa-Kawada, J., Suzuki, R., Kano, F., Ohkawara, T., Murata, M. and Noda, M. (2003) Axonal morphogenesis controlled by antagonistic roles of two CRMP subtypes in microtubule organization. Eur. J. Nuerosci., 17, 2329-2343. Ohyama, K., Ikeda, E., Kawamura, K., Maeda, N. and Noda, M. (2004) Receptor-like protein tyrosine phosphatase z/RPTPb is expressed on tangentially aligned neurons in early mouse neocortex. Develop. Brain Res., 59, 34-47. Shintani, T., Kato, A., Yuasa-Kawada, J., Sakuta, H., Takahashi, M., Suzuki, R., Ohkawara, T., Takahashi, H. and Noda, M. (2004) Large-scale identification and characterization of genes with asymmetric expression patterns in the developing chick retina. J. Neurobiol., 148, 121-127. |