RESEARCH CENTER FOR INTEGRATIVE AND COMPUTATIONAL BIOLOGY
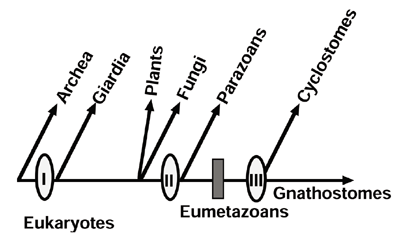
The neutral theory of molecular evolution, the basic theory for explaining evolutionary changes at the molecular level, claims that most evolutionary changes at the molecular level are caused by random drift of neutral mutants. This is sharply contrasted to the evolution at the organismal level, where it is generally thought that natural selection is the major driving force by which evolutionary changes occur. How can we explain the evolutionary changes at the two levels in a unified way. This is the major problem that remains unsolved for molecular evolutionists. As a first step for understanding the final problem, we are investigating relationships of evolutionary diversifications between molecular and organismal levels, based on molecular phylogenetic approach. I. Divergence pattern of animal gene families and relationship with the Cambrian explosionIn multicellular animals, a variety of gene families involved in cell-cell communication and developmental control have evolved through gene duplication and gene shuffling, basic mechanisms for generating diverse genes with novel functions. Each of these animal gene families has originated either from a few ancestral genes which are shared with plants and fungi or from an ancestral gene created uniquely in animal lineage. The major groups of bilateral animals are thought to have diverged explosively at or prior to the Vendian - Cambrian boundary. No direct molecular evidence has been provided to date as to whether the Cambrian explosion was triggered by a dramatical increase in the number of genes involved in cell-cell communication and developmental control either immediately prior to or in concert with the Cambrian explosion. A molecular phylogeny-based analysis of several animal-specific gene families has revealed the gene diversification by gene duplication occurred during two active periods interrupted by a long intervening quiescent period. Intriguingly, the Cambrian explosion is situated in the silent period, indicating that there is no direct link between the first burst of gene diversification and the Cambrian explosion itself. The above result also suggests the importance of gene recruitment as a possible molecular mechanism for morphological diversity. II. Episodic divergence of eukaryote-specific genes and evolution of Giardial membrane bounded organellaTo understand a question of whether divergence of eukaryote-specific gene families recognized commonly in all eukaryotes by gene duplication and domain shuffling proceeded intermittently during evolution, as found in animal-specific gene families, a further molecular phylogeny-based analysis has been conducted for kinesin family as a typical example for eukaryote-specific gene families. We have cloned and sequenced cDNAs encoding kinesins and kinesin-related proteins (KRPs) Giardia lamblia, the most primitive protist that represents the earliest branching among extant eukaryotes and have obtained 13 kinesin-related cDNAs, some of which are likely homologs of vertebrate kinesins involved in vesicle transfer to ER, Golgi, and plasma membrane. A phylogenetic tree of the kinesin family revealed that most gene duplications that gave rise to different kinesin subfamilies with distinct functions have been completed before the earliest divergence of extant eukaryotes. This suggests that the complex endomembrane system has arisen very early in eukaryotic evolution, and diminutive ER and Golgi apparatus recognized in the Girdial cells, together with the absence of mitochondria, might be characters acquired secondarily during evolution of parasitism. To understand the divergence pattern of the kinesin family in the lineage leading to vertebrates, seven more Unc104-related cDNAs have been cloned from sponge, amphioxus, hagfish, and lamprey. The divergence pattern of animal Unc104/KIF1 subfamily is characterized by two active periods in gene duplication interrupted by silent periods of considerable long, but not proceeded gradually: Thus, in the eukaryotic lineage leading to vertebrates, the divergence of kinesin family might occurred episodically, as demonstrated by at least three periods of extensive gene duplications, instead of proceeding gradually. Extensive subfamily-generating duplications in ancient times before the earliest branching among extant eukaryotes may have implications to the evolution of Giardial membrane bounded organella.
Publication ListInokuchi R., Kuma, K., Miyata, T. and Okada, M. (2002) Nitrogen-assimilating enzymes in land plants and algae: Phylogenetic and physiological perspectives. Physiologia Plantarum 116, 1-11. Iwabe, N. and Miyata, T. (2002) Kinesin-Related Genes from Diplomonad, Sponge, Amphioxus and Cyclostomes: Divergence Pattern of Kinesin Family and Evolution of Giardial Membrane Bounded Organella. Mol. Biol. Evol. 19, 1524-1533. Katoh, K., Misawa, K., Kuma, K. and Miyata, T. (2002) MAFFT: a novel method for rapid multiple sequence alignment based on fast Fourier transformation. Nucleic Acids Res 30, 3059-3066. Katoh, K. and Miyata,T. (2002) Cyclostome hemoglobins are possibly paralogous to gnathostome hemoglobins.J. Mol. Evol. 55, 246-249.
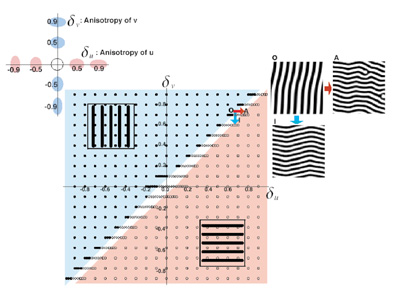
I. Directionality of Stripes Formed by Anisotropic Reaction-Diffusion Models.The pattern formation of animal coating has been studied mathematically by a pair of partial differential equations, named a reaction-diffusion (RD) model. By the model, starting from an initial distribution very close to uniformity, a spatial heterogeneity emerges and a stable periodic pattern is formed spontaneously. This simple mechanism suggests that the reaction of a small number of chemicals and their diffusion might create stable non-uniform patterns. When we analyze the model in a two dimensional plane, striped patterns in addition to spotted patterns often emerge. This was considered as the basic mechanism explaining the stripe patterns observed among animal coating. We focused on the directionality of the stripes. Most of the stripes observed in the fish skins are either parallel or perpendicular to their anterior-posterior (AP) axis. The direction of stripes is considered of importance in the behavioral and ecological viewpoints. However, very little is known about the mechanisms that makes the strong directionality either in the actual fish skin or in the theoretical models. The standard RD model doesn't determine the direction of stripes. To explain the directionality of stripes on fish skin in closely related species, we have studied the effect of anisotropic diffusion of the two substances on the direction of stripes, in the cases in which both substances have the high diffusivity in the same direction. We also studied the direction of stripes in more general situations in which the diffusive direction may differ between the two substances. We derived a formula for the direction of stripes, based on a heuristic argument of unstable modes of deviation from the uniform steady state. We confirm the accuracy of the formula by computer simulations. When the diffusive direction is different between two substances, the directions of stripes in the spatial pattern change smoothly with the magnitude of anisotropy of two substances. When the diffusive direction of the two substances is the same, the stripes are formed either parallel to or perpendicular to the common diffusive direction, depending on the relative magnitude of the anisotropy. The transition between these two phases occurs sharply.
II. Comparative Study of Circadian Clock ModelsMany species living in an environment fluctuating with a strong daily rhythm have evolved an internal clock. According to molecular biological studies of circadian clock, many genes and proteins are involved in the biochemical dynamics generating their stable rhythm. Although the identity of genes and proteins may differ between species, the basic mechanism is the same -- there is one or more genes whose products may enter to the nucleus and then suppress the transcription of their own gene(s). This negative feedback with a long time delay can generate the circadian rhythm. In this study, we attempt to identify those aspects of biochemical systems for the circadian rhythm which promote sustainable oscillation. We discussed models of different complexity, and examined the effects of cell compartmentalization, protein modification steps in chemical reactions, having multiple clock genes in a cell, and cooperativity in the inhibition of transcription and in the protein transport. To derive general biological conclusions, we focused on mathematical analyses of relatively simple models, rather than computer simulations of complex realistic models. First, using a Lyapunov function, we proved under very general conditions that two-variable models always converge to a stable equilibrium, implying that additional structures are needed for the model to generate a sustainable rhythm. Second, we compared several models of different complexity using the Routh-Hurwitz criteria of stability. We showed that a sustainable oscillation is more likely to occur; if the cell is compartmentalized so that the proteins need to be transported from the cytosol to the nucleus; if the proteins have to be modified before entering the nucleus; if the kinetics of transcription inhibition or the transport to the nucleus have a nonlinear dependence on the substrate concentration (cooperativity); or if the products of two clock genes form a heterodimer that suppresses both of their own genes. Specifically we analyzed the local stability mathematically for the cases: (1) there is the reaction reverse to protein modification, (2) there are two clock genes, per and tim, instead of one -- which have been analyzed only by computer simulations; and also the cases (3) the protein transport to the nucleus has cooperativity, which has not been examined before. III. Pattern formation of the cone mosaic in the zebrafish retinaIn teleost fish, there are several subtypes of cone cells, which are sensitive to different wave-lengths of light. In retinas of some species of teleost fish, regular arrangements of cone cells are observed, where each subtype of cone cells appears periodically in the two-dimensional retinal sheet. Different patterns are observed in different species. These patterns are called "cone mosaics." The biological mechanism of the pattern formation is still under examination. Some species show quite different mosaic patterns between peripheral region and more central region in the retina, which suggests mobility of cone cells in retinal space. We have studied the mechanism of the pattern formation by using mathematical models. In this study, I examined a process of cell-cell interaction to generate the regular mosaic pattern -- namely cell rearrangement. In the model, cells have already been determined as to their final subtypes, but they change their locations in the pattern formation process and the cell movement is affected by their neighbours. I study the conditions for generating mosaic pattern. First, an assumption regarding "double cone" is shown to be necessary, whereby the two component cells of a double cone behave as if they are a single cone, i.e. a green cone and a red cone are never detached during the pattern formation process. Second, the correct pattern is derived by computer simulation only when the chosen magnitude of cell adhesion is appropriate. I determine the conditions of adhesive strength for generating the observed pattern. These conditions lead to the testable predictions.
I also compared the condition of cell-cell interaction for generating the mosaic pattern with the different model, the fate transition model. In the fate transition model, cell differentiation regulated by interaction of neighbouring cells is assumed to be responsible for the mosaic pattern. The condition for obtaining the focal pattern is looser in the cell rearrangement model than in the fate transition model.
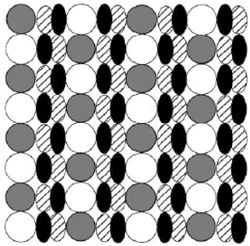
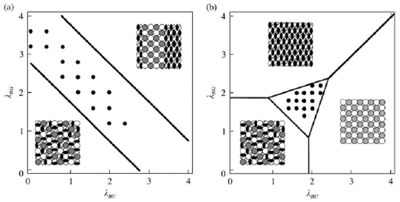
IV. Formation and Maintenance of Distinctive Cell Patterns by Co-expression of Membrane-bound Ligands and Their ReceptorsWe showed that graded or checkerboard-like cell patterns, and segmental domains along a body axis, can be generated by cell behaviors involving differences in intercellular repulsion. A membrane-bound signal transduction system mediating contact-dependent cell interactions includes membrane-bound ligands (ephrins) and their receptors with tyrosine-kinase activity (Eph proteins). These molecules mediate both repulsive and attractive interactions under bilateral threshold control: Cells expressing the receptors adhere to a surface bearing a critical density of ligand reciprocal to the density of receptor, but are repelled by a surface with other densities of ligand. The present paper extends this model: General membrane-bound ligands (not always ephrins) and their receptors are assumed to be co-expressed in a single cell under bilateral threshold control. Computer simulations of cell pattern formation showed that when co-expression of the ligand and receptor is reciprocal, the cells self-organize into a pattern of segmental domains or a graded cell arrangement along the body axis. The latter process interprets positional information in terms of protein molecules. When co-expression of the two species of molecules is not always reciprocal, the cells generate various patterns including checkerboard and kagome (star) patterns. The case of separate expression of ligands and receptors in different cells is also examined. The mechanism of differences in cell repulsion is compared to the differential cell adhesion hypothesis, which has been used to explain cell sorting. Publication ListHonda, H. and Mochizuki, A. (2002) Formation and maintenance of distinctive cell patterns by co-expression of membrane-bound ligands and their receptors. Dev. Dyn. 223, 180-192. Kurosawa, G., Mochizuki, A. and Iwasa, Y. (2002) Processes promoting oscillations -- comparative study of circadian clock models. J. Theor. Biol. 216, 193-208. Mochizuki, A. (2002) Pattern formation of cone mosaic in zebrafish retina: A Cell rearrangement model. J. Theor. Biol. 215, 345-361. Shoji, H., Mochizuki, A., Iwasa, Y. and Kondo, S. (2002) Directionality of Stripes Formed by Anisotropic Reaction-Diffusion Models. J. Theor. Biol. 214, 549-561. Tohya, S., Mochizuki, A. and Iwasa, Y. Random cell sorting can form cone mosaic patterns in fish retina and explain the difference between zebrafish and medaka. J. Theor. Biol. (In Press) Ryohji, T., Mochizuki, A. & Iwasa, Y. Possibility of Tissue Separation Caused by Cell Adhesion. J. Theor. Biol.(In Press) |
|
Copyright(C) National Institute for Basic Biology (webmaster@nibb.ac.jp) |