DIVISION OF SPECIATION MECHANISMS I
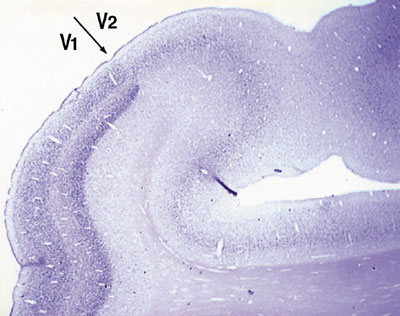
Our research is focusing to understand mechanisms underlying memory and evolution of the brain. For one approach to understand these questions, we are studying the genes that are expressed in specific areas of the primate neocortex. Using differential display method, we obtained three genes that showed marked differences within areas of the primate neocortex. Our second approach is to understand informational processing in the brain underlying learning behaviors by examining gene expression. Here, we report our findings focusing on those of the year of 2002. I. Genes expressed in specific areas of the neocortexThe neocortex, most remarkably evolved in primates, plays the major role in higher functions of the brain. It is known to be divided into distinct functional and anatomical areas and has been a matter of debate to what extent the area of the neocortex is genetically and environmentally determined. It is also puzzling why, during the evolution of mammals, the neocortex was most markedly expanded while the number of the genes in mammal phyla were little changed. To access these questions, we studied gene expression within different areas of the neocortex. 1) In collaboration with Professor Hiroyuki Nawa (Nigata university), we used the DNA macroarray technique to examine gene expression in the areas of human prefrontal, motor and visual cortexes. We found almost all the genes among 1088 genes examined showed only less than a factor of two in the difference of their expressions. Only one gene showed more than three fold difference and another one was between two and three fold difference within the three areas. These results suggest that the genes that are expressed among different areas of the human neocortex are very similar. However, the question remained whether there are any genes that show marked differences within areas of the neo-cortex. 2) In order to answer this question, we employed differential display methods and found at least three genes that indicated area specific expressions. i) One gene, designated occ1, is specifically expressed in the occipital cortex, particularly in V1, in the primate brain. Furthermore, the expression of occ1 turned out to be activity dependent, because, in the monocularly deprived monkeys injected with TTX into one of the eyes, the expression of occ1 is markedly decreased in the ocular dominance column of the primary visual cortex (V1). ii) The other gene that showed marked difference within the neocortex, is gdf7, a member of BMP/TGF-b family, which is specifically expressed in the motor cortex of African green monkeys. We are currently examining the detailed expression pattern of the gene. iii) The third gene that we designated tentatively as 134G was preferentially expressed in association and higher sensory areas in the neocortex (Komatsu et al., Society for Neuroscience 32nd Annual Meeting, 2002). 3) We have also further isolated several area specific genes with RLCS (Restriction Landmark cDNA Scanning). We are now characterizing these genes to reveal the mechanisms that form neocortical areas.
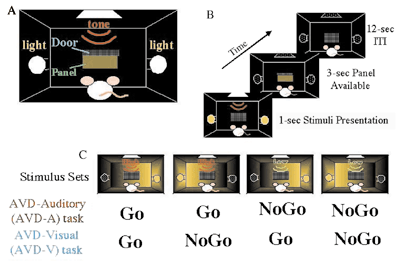
II. Gene expression under a declarative and a non-declarative memoryIn order to study informational processing underlying the declarative and the non-declarative memory at molecular and cellular levels in the brain, we employed c-Fos mapping techniques. There have been an increasing number of studies using c-Fos as markers to examine neuronal activities since c-Fos induction by electrical stimulation was found by Morgan and Curran. However, many sensory stimuli per se are now known to cause c-Fos induction. So, we should be very careful to distinguish the c-Fos expression that is caused by learning process from that caused by sensory stimuli. For this purpose, it is necessary to use behavioral systems that are able to distinguish the two. Although a few behavioral systems in rodents have been successfully used for physiology, animal behavior and recently for analyses of knockout mice, little behavioral systems in fact distinguish them. Therefore, we prepared ourselves for using two behavioral systems, each of which represent a declarative and a non-declarative memory. In one system, we collaborated with professor Yoshio Sakurai (Kyoto University) who developed audio-visual discrimination task (AVD-task). In this task, a rat was asked to choose either an audio cue (a high or low tone) or a visual cue (a light from the right or the left) to obtain a food pellet (Fig. 2). We found that the visual and audio tasks enhanced the specific expression of c-Fos in the visual and audio cortices, respectively. Among the early visual and auditory pathways examined, c-Fos was specifically induced in the cortices but not in the earlier pathways, suggesting the neural modulation of the neocortex depending on the types of the tasks. Interestingly, the task-dependent c-Fos expression was only observed in excitatory neurons in the relevant sensory cortices.
The other task we developed is a wheel running system in which a water-deprived mouse is asked to run to obtain water in front because the wheel with pegs is turning to the other direction (Kitsukawa et al., Society for Neuroscience 32nd Annual Meeting, 2002). The pegs can be changed with various patterns as desired. The task required for the mouse thus can be regarded to represent a procedural learning. We examined various areas of brains following to the change of the peg pattern. Among the areas examined, we found marked c-Fos expression in the striatum. The striatum, which is composed of projection neurons and several distinguished types of interneurons, is known to play an important role in a reward-based learning. The characterization of these subtypes of interneurons has been progressed. However, the roles in behavioral tasks have been little known. We hope our system combined with c-Fos mapping technique solve these questions. III CNTF is specifically expressed in the developing pineal glandCNTF, a member of the IL-6 family, attracts quite attentions of developmental neuroscientists because it shows various effects on neurons and glias. CNTF knockout mice, however, only indicate moderate motor neuron deficiency in the adult, but no apparent phenotype in the development. In order to explore the function of the IL-6 family, we extensively examined the expression of members of the family and their receptors and found the specific expression of CNTF in the embryonic pineal glands and eyes. This was to our knowledge for the first time to show a clear expression of CNTF in the development. Next question to be asked is the functional role of CNTF in the pineal development. Because of no reported phenotype of CNTF knockout mice in their development, it has been difficult to know the developmental roles of CNTF if any. In fact, we confirmed that there seemed no apparent difference in the pineal development of CNTF knockout mice compared to that in wild type mice. This is presumably because the CNTF gene knockout was compensated by other CNTF-like factors. We therefore studied cultured pineal organs in rodents to ask if there were any effects of CNTF on them following to a previous study that shows that cultured neonatal pineal organs develop photoreceptor-like cells (Araki, 1992). We found that CNTF inhibits photoreceptor-like cells (Hata et al., Society for Neuroscience 32nd Annual Meeting, 2002). Our observation raises an interesting possibility that CNTF plays a critical role in the pineal development to suppress a certain phenotype such as photoreceptors. We are currently working to prove it. Publication list:Hata, K., Arakai, M. and Yamamori, T. (2002) CNTF is specifically expressed in the developing rat pineal gland and eyes. NeuroReport 13, 735-739 Nakayana, T., Mikoshiba, K., Yamamori, T. and Akazawa, K. Expression of syntaxin 1C, an alternive splice variant of HPC-1/syntaxin 1A, is activaed by phorbol-ester stimulaton in astrglioma. FEBS Lett. (in press) Onishi, A., Koike, S., Ida-Hosonuma, M., Imai, H., Shichida, Y., Takenaka, O., Hanazawa, A., Komatsu, H., Mikamai, A., Goto., S., Suryobroto, B., Farajallah, A., Varavudhi, P., Ekavhibata, C., Kitahara, K. and Yamamori, T. (2002) Variations in ling-term and middle-wavelength-sensitive opsin loci in crab-eating monkeys. Vision Res., 42, 281-292 Sakata, S., Kitsukawa, T., Kaneko, T., Yamamori, T. and Sakurai, Y. (2002) Task-dependent and cell-type-specific Fos enhancement in rat sensory cortices during audio-visual discrimination. Eur. J. Neuroscience.15, 735-743 Suga, K., Yamamori, T. and Akagawa, K. Identification of Carboxyl-Terminal Membrane-Anchoring Region of HPC-1/Syntaxin 1A by Substituted-Cysteine-Accessibility Method and Monoclonal Antibody. J. Biolchem (in press) Tochitani, S., Hashikawa, T. and Yamamori, T. Expression of occ1 mRNA in the visual cortex during postnatal development in macaques. Neurosci. Lett. (in press) Vigot, R., Batini, C., Kado, R.T. and Yamamori. T. (2002) Synaptic LTD in vivo recorded on the rat cerebellar cortex. Arch. Ital. Biol., 140, 1-12. |
|
Copyright(C) National Institute for Basic Biology (webmaster@nibb.ac.jp) |