DIVISION OF GENE EXPRESSION AND REGULATION II
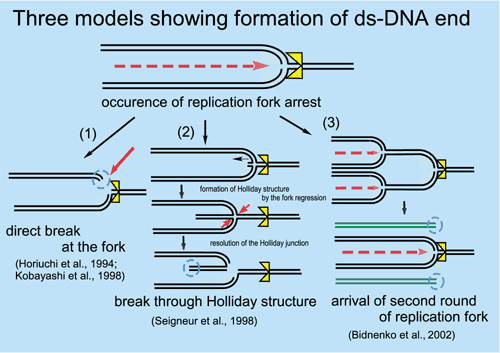
* to Biomolecular Engineering Research Institute, 6-2-3, Furuedai, Suita, Osaka, 565-0874, JAPAN (Sep, 1, 2002- ). The genomes of higher organisms contain significant amounts of repetitive sequences, which in general, are unstable. At present, neither the physiological function(s) of repeated sequences nor the mechanism controlling the instability is fully understood. To clarify these aspects, we are pursuing the following themes using E. coli and S cerevisiae: (1) An amplification mechanism of repeated sequences or genes, especially rRNA repeated genes, (2) a mechanism of replication fork block-dependent recombination, a key reaction that increases or decreases the number of repeats, and (3) development of in vivo artificial gene amplification systems. Structural and functional analyses of the E. coli genome are also being carried out. In 2002, work on the following three subjects has advanced our knowledge of the dynamics and structure of the genome. I. Unequal sister-chromatid recombination induces the instability of the rDNA in Saccharomyces cerevisiae.The yeast S. cerevisiae carries approximately 150 copies of the ribosomal RNA gene (rDNA) in tandem repeats on chromosome XII. Transcription of the rDNA is carried out specifically by RNA polymerase I (Pol I), one of the three kinds of eukaryotic RNA polymerases. The absence of an essential subunit of Pol I in rpa135 deletion mutants causes a gradual decrease in the rDNA repeat number to approximately half the normal level and reintroduction of the missing RPA135 gene induces a gradual increase in repeat number back to the normal level. This contraction/expansion requires the FOB1 gene protein (Fob1p) which is also required for replication fork blocking activity at the replication fork barrier (RFB) site. The RFB site is located in a non-transcribed spacer region, called NTS1, in each rDNA unit. Using this artificial induction system of amplification, we tested the effects of several mutations on the pattern of copy number increase. The gene RAD52 was essential for the increase in rDNA repeats. mre11 mutant cells showed a partial reduction in the amplification of rDNA. In a sir2 mutant, the rate of amplification increased by more than two-fold, although the copy number was not stabilized. The hyper-recombinogenic phenotype in the mutant was dependent on FOB1. In addition, the loss of SIR2 function reduced the association of Mcd1p, a factor involved in sister-chromatid cohesion, with the non-transcribed spacer region, NTS1, in the rDNA repeats. A temperature sensitive mutant of the cohesin complex, smc1-2, had an approximately 8-fold increase in the loss of a URA3 marker, which is integrated in the rDNA repeats, at the semi-permissive temperature. These results suggest that the establishment of sister-chromatid cohesion would affect the efficiency of the unequal sister-chromatid recombination to change the copy number of rDNA after the double strand break at the DNA replication fork arrested at the RFB site (see Figure). II. Transcription-dependent recombination in yeast rDNA.In Escherichia coli, we previously identified a chromosome-derived EcoRI fragment, as a recombinational hotspot, HotH, which contains the 3' end of an rRNA operon, rrnD. Under HotH active conditions, the progress of the DNA replication fork was retarded in the latter part of the rrnD operon, the transcriptional orientation of which is opposite to that of the fork. This result suggests that the collision between the replication fork and rRNA gene transcription may be responsible for retardation. Inactivation of the promoter of the rrnD operon simultaneously reduced the retardation of fork movement and HotH activity. These results indicate that HotH activation is caused by the collision between the DNA replication fork and the reverse-oriented rrnD transcription. The rRNA gene (rDNA) in S. cerevisiae has a tandem repeat structure, whose copy number is approximately 150 copies. The replication fork barrier (RFB) site is located near the 3'-end of the 35S rDNA of each rDNA unit. This site inhibits the progression of the DNA replication fork coming from the opposite direction of the 35S rDNA transcription. The gene FOB1 was previously shown to be required for the replication fork blocking activity at the RFB site and also later for the change in the copy number of rDNA through unequal sister chromatid recombination. Another function of the RFB site may be to avoid a collision between 35S rDNA transcription and DNA replication. Although a collision has never been observed in the wild type or even in a fob1 mutant with 150 copies of rDNA, it could be detected in a fob1 derivative with a reduced number of rDNA copies (approximately 20) by using two-dimensional agarose gel electrophoresis. This result suggests that while each rDNA of 150 copies may be weakly transcribed or a small fraction of the rDNA may be actively transcribed, most of the reduced copies are probably extensively transcribed. The collision was dependent on the transcription of 35S rDNA by Pol I. In addition, under these FOB1 defective conditions, transcription apparently enhanced recombination to change the copy number of rDNA and also to produce the extra-chromosomal rDNA circles (ERCs) whose accumulation is known to be a cause of aging. These results suggest that such a transcription-dependent collision may function as a recombination trigger, such as introducing a ds-break shown in Figure, for multiplication of a single copy gene that is intensely transcribed in general.
III. Complete genomic sequence of the E. coli K12 W3110 strainEscherichia coli is one of the organisms that has been most extensively analyzed physiologically, biochemically and genetically. Of all E. coli strains, E. coli K12 W3110 has probably been used most frequently as the wild-type strain in these experiments. Here, we have determined the complete nucleotide sequence of the genome of strain W3110 by mainly using lambda phages of Kohara's bank. Previously a US group determined the genomic sequence of another K12 wild-type derivative, MG1655. Both strains were derived from a common ancestor strain, W1485, approximately 50 years ago. After comparing the two sequences , the following results were obtained. The total number of nucleotides in the W3110 genome is 4660170 bp. There were 349 bp conflicts between the sequences of W3110 and MG1655. Re-sequencing of the conflict sites by a PCR method using their genomic DNAs as templates revealed that only eight sites (9 bp) were true conflicts. Seven of them are base-change type conflicts and one is a two base frame-shift. All of these differences reside within genes, seven in ORFs and one in a 23S rRNA gene. Next we investigated the number and the type of insertion sequence (IS) elements in collaboration with Dr. Ohtsubo's laboratory (Tokyo Univ.). Thirteen different IS elements were identified. While 12 IS elements are present in the W3110 genome and not in MG1655, one IS element is in MG1655, but not in W3110. These results indicate that the DNA sequence is almost perfectly conserved in bacterial cells under stock culture conditions and in Kohara's lambda phage bank, and during growth through repeated vegetative cycles10. Under stock conditions, which include transfers from the stock to fresh plates and vice versa, IS transposition events seem to occur more frequently than base-changes. Publication list:Johzuka, K., and Horiuchi, T. (2002) Replication fork block protein, Fob1, acts as an rDNA region specific recombinator in S. cerevisiae. Genes Cells 7:99-113. Kodama, K., Kobayashi, T., Niki, H., Hiraga, S., Ohshima, T., Mori, and Horiuchi, T. (2002) Amplification of HotDNA segments in Escherichia coli. Mol. Microbiol. 45: 1575-1588. |
|
Copyright(C) National Institute for Basic Biology (webmaster@nibb.ac.jp) |