DIVISION OF GENE EXPRESSION AND REGULATION I
1)from Kasetsart University The main interest of the group is in understanding the biology of the dynamic genome, namely, genome organization and reorganization and its impact on gene expression and regulation. Although there are many elements affecting organization and reorganization of the genomes, we are currently focused on mobile genetic elements in general and plant transposable controlling elements in particular. Since plant transposable elements are known to be associated with both genetic and epigenetic gene regulations, we are characterizing various aspects of genetic and epigenetic gene regulations. In addition, we are also undertaking reverse genetic approaches in order to elucidate the nature of dynamic genome in plants. I. Spontaneous mutants in the Japanese morning glory.The Japanese morning glory (Ipomoea nil or Pharbitis nil), displaying blue flowers, is believed to be originated from southeast Asia and has an extensive history of genetic and physiological studies. The plant had been introduced into Japan from China in about 8th century as a medicinal herb, seeds of which were utilized as a laxative, and has become a traditional horticultural plant in Japan since around 17th century. A number of its spontaneous mutants related to the color and shape of the flowers and leaves have been isolated, and about 10% of these mutants carry mutable alleles conferring variegated phenotypes. All of the mutants available are spontaneous mutants and most of them were isolated more than 60 years ago. Several lines of evidence indicate that an En/Spm-related transposable element Tpn1 and its relatives, which we termed Tpn1-family elements, are common spontaneous mutagens in the plant. Indeed, we previously identified four mutable alleles for flower pigmentation, flecked-1, flecked-2, speckled and purple-mutable (pr-m), which are caused by integration of non-autonomous Tpn1-related elements, Tpn1, Tpn9, Tpn2 and Tpn4 into the genes for antocyanin biosynthesis, DFR-B, ANS, CHI and the gene for a vacuolar Na+/H+ exchanger InNHX1, respectively. An apparent stable r-1 allele conferring white flowers was also previously found to be caused by insertion of a non-autonomous Tpn1-family element, Tpn3, into the CHS-D gene encoding a chalcone synthase for anthocyanin biosynthesis. The transposition of these non-autonomous elements is mediated by a Tpn1-related autonomous element, which appears to be subjected to epigenetic regulations. Thus the phenotype of the white flowers in the r-1 mutant due to very rare excision of Tpn3 from the CHS-D gene is likely to be the result from epigenetic inactivation of the autonomous element. We are able to identify two mutations in the UF3GT gene encoding UDP-glucose:flavonoid 3-O-glucosyltrans-ferase, which is the first step to form stable anthocyanin pigments accumulating in the vacuole. One of them carries a Tpn1-related non-autonomous element Tpn10 having integrated into its promoter region, and the other contains an insertion of 4 bp, which generates a termination codon in the cording region. Another mutant displaying maroon flowers was found to carry a 4-bp insertion within the coding region of a newly isolated glucosyltransferase gene. The mutant is likely to be deficient in a transglucosylation process from anthocyanidin 3-glucose to the poly-acylated and glucosylated anthocyanin pigments accumulated in the petal vacuole. In addition to these newly identified mutations in the genes encoding the enzymes involved in anthocyanin biosynthesis pathway, mutants bearing white flowers were found to carry two different 7 bp insertions at the same site within the coding region of a regulatory protein containing the WD40 motif. Both insertions causing frameshifts would result in truncated regulatory protein. These 7 bp insertions are likely to be footprints generated by excision of a Tpn1-related transposable element. The results imply that 4 bp insertions detected in the two different glucosyltransferase genes are also footprints generated by excision of Tpn1-related elements, which are spontaneous mutagens in the Japanese morning glory.
II. Spontaneous mutants in the common morning glory.The common morning glory (Ipomoea purpurea or Pharbitis purpurea), originating from Central America, was introduced to Europe probably in the late 17th century and several cultivars with different color of flowers were already recorded in the late 18th century. Previously, we identified that the mutable flaked allele of the plant for flower variegation is caused by the insertion of the transposable element Tip100 into the CHS-D gene. The 3.9-kb Tip100 element belongs to the Ac/Ds family. We are able to identify two mutant lines showing white seeds and palely pigmented flowers were found to produce drastically reduced amounts of the transcripts of the IPMYC3 gene, a homologue of the an1 gene for the bHLH transcriptional factor that regulates flower and seed pigmentation in petunia. The genomic sequences revealed that both mutants carry two insertions of transposable elements into the IPMYC3 gene, which comprises 8 exons. In one mutant, two copies of Tip100 belonging to the Ac/Ds family are inserted into the 2nd intron and the 7th exon. The other mutant contains two different Mu-related transposable elements in the 2nd exon and the 5th intron. Moreover, the latter mutant appears to carry an additional mutation for flower pigmentation. III. Spontaneous mutants in the morning glory (Ipomoea tricolor).Like Ipomoea purpurea, the morning glory (Ipomoea tricolor) is also originated from central America. The wild type plant produces bright blue flowers, which is one of the most important floricultural plants in both the United States and Europe. We have identified a mutant displaying white flowers carries about 11.5 kb insertion into the 5th intron of the DFR-B gene. The junction sequences of the insertion site revealed that the 5th intron of the DFR-B gene contains a 0.7 kb MITE-like element and that the 11.5 kb sequence is integrated in the middle of the element. Neither terminal inverted repeats of the insertion nor its target site duplication was detected, and the transcription of the DFR-B gene appears to be blocked efficiently within the inserted sequence.
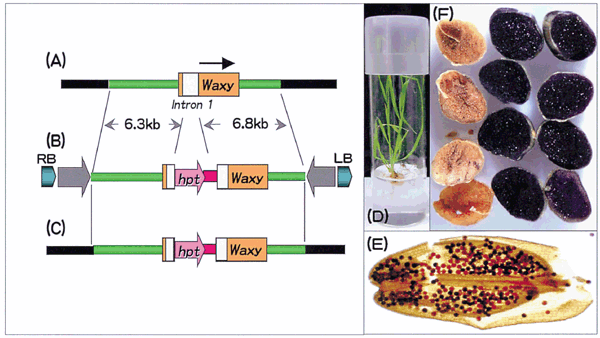
IV. Targeted gene disruption by homologous recombination in rice.Rice (Oryza sativa L.) is an important staple food for more than half of the world's population. It is a model plant for other cereal species because of several characteristics: its small genome of about 430 Mb; the similarity of its sequences and constellation of the genes with other cereals; the availability of large expression sequenced tag (EST), full-length cDNAs and the entire genome sequence of both Japonica and Indica subspecies; and its efficient use of Agrobacterium-mediated transformation. A comparison of the genome sequences of these Japonica and Indica subspecies with that of Arabidopsis indicates that a large proportion of rice genes hove no recognizable homologues in Arabidopsis. Under these circumstances, the development of a method to study gene function by modifying genomic sequences precisely becomes extremely important, and gene targeting is a powerful tool of such reverse genetics. In higher plants, however, the method is far from a common practice. We have succeeded to develop an efficient and reproducible procedure for gene targeting in rice, based upon the following components: (a) optimization of Agrobacterium-mediated transformation of embryogenic rice calli, (b) utilization of strong positive/negative selection, and (c) stringent PCR screening for targeted allele. As a model gene to be targeted, we chose the Waxy gene encoding granule-bound starch synthase, a key enzyme in amylose synthesis, because its mutations affect the quality and quantity of rice grain and because the associated phenotype in pollen and in endosperm can easily assessed by simple iodine staining. All of the six independently obtained fertile transgenic rice plants from six experiments were heterozygotes at the Waxy locus: a wild-type Waxy allele and a targeted recombinant waxy allele. These transgenic plants segregated into a 3:1 ratio, confirming to the Mendelian pattern of inheritance. Neither ectopic targeting (integration of the sequence produced by homologous recombination into a site other than the correctly targeted site in the genome) nor ectopic integration (integration by nonhomologous endjoining) of the selective marker gene could be observed. Southern blot analysis revealed that the targeted waxy region of around 35 kb comprised the anticipated structure and that the drug-resistant marker used for selecting transformants was integrated only within the targeted waxy gene in the genome. Sequencing analysis of the junction regions confirmed that precise somatic homologous recombination occurred to generate the rice plants carrying the targeted waxy gene. We are currently examining whether the strategy we used is applicable to obtain various gene-targeted or knockout lines of rice. V. Characterization of mutable virescent allele in rice.Leaves of seedlings in the virescent mutant of rice (Oryza sativa L.) are initially pale yellow green due to partial deficient in chlorophyll and gradually become green with the growth of the mutant. The mutable virescent plant (yl-v), displaying pale green leaves with dark green sectors, was isolated among progeny of a hybrid between Indica and Japonica rice plants. The leaf variegation is regarded as a recurrent somatic mutation from the recessive pale green to the pigmented revertant allele. During repeated crossing with the Japonica rice T-65, an apparent stable virescent mutant, yl-stb, the seedlings of which bear pale green leaves, was obtained. Although the yl-stb mutant could not grow on soil, we have succeeded in growing it with in vitro culture during its critical juvenile phase. Preliminary HPLC analysis indicated accumulation of a carotenoid intermediate in the yl-stb mutant. The mutable virescent allele was mapped in the short arm of the rice chromosome 3.
Publication List:Iida, S. and Terada, R. (2002) Gene modification of an endogeneous gene in rice plants. ISB News Report December 2002, 7-8. Ishikawa, N., Johzuka-Hisatomi, Y., Sugita, K., Ebinuma H. and Iida, S. (2002) The transposon Tip100 from the common morning glory is an autonomous element that can transpose in tobacco plants. Mol. Gen. Genomics 266, 732-739. Kamiunten, H., Inoue, S., Yakabe Y. and Iida, S. (2002) Characterization of ISPsy2 and ISPsy3, newly identified insertion sequences in Pseudomonas syringae pv. Eriobotryae. J. Gen. Plant Pathol. 68, 75-80. Terada, R., Urawa, H., Inagaki, Y., Tsugane, K. and Iida, S. (2002) Efficient gene targeting by homologous recombination in rice. Nature Biotechnol. 20, 1030-1034. |
|
Copyright(C) National Institute for Basic Biology (webmaster@nibb.ac.jp) |