DIVISION OF BIOENERGETICS
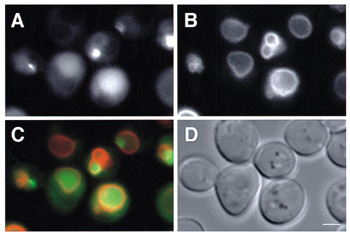
I. BackgroundThis division aims to understand the autophagy in respects of its molecular mechanism and its physiological role within higher eukaryotes. Biosynthetic processes and degradation processes are well coordinated to regulate the biological activities, however the study on the latter has been retarded compared to the one on the former and we must shed light on the degradation process to fully understand the cell. Autophagy is a well conserved degradation process in eukaryotes and is a major route for bulk degradation of cytoplasmic constituents in a lytic compartment, lysosome/vacuole. Upon nutrient starvation, autophagic process starts in the cytoplasm as building up of a membrane structure, an autophagosome, which sequesters a portion of cytosol. Then it is delivered to the vacuole/lysosome, and the cytoplasmic materials inside are degraded by the vacuolar/lysosomal proteases. We had discovered autophagy in a simple eukaryotic model organism, Saccharomyces cerevisiae and morphologically described its whole process. We have isolated a set of autophagy-deficient mutants (apg), and cloned APG genes essential for autophagy. 2. Formation of the Apg12-Apg5-Apg16 multimeric complex is essential for autophagy in yeast.Autophagy requires a ubiquitin-like protein conjugation system, in which Apg12 is covalently bound to Apg5. In the yeast Saccharomyces cerevisiae, the Apg12-Apg5 conjugate further interacts with a small coiled-coil protein, Apg16. The Apg12-Apg5 and Apg16 are localized in the cytosol and pre-autophagosomal structures and play an essential role in autophagosome formation. We showed that the Apg12-Apg5 conjugate and Apg16 form an approximately 350-kDa complex in the cytosol. Because Apg16 was suggested to form a homo-oligomer, we generated an in vivo system that allowed us to control the oligomerization state of Apg16. With this system, we demonstrated that formation of the approximately 350-kDa complex and autophagic activity depended on the oligomerization state of Apg16. These results suggest that the Apg12-Apg5 conjugate and Apg16 form a multimeric complex mediated by the Apg16 homo-oligomer, and formation of the approximately 350-kDa complex is required for autophagy in yeast. 3. Cargo Delivery to the Vacuole Mediated by AutophagosomesIn the yeast Saccharomyces cerevisiae, aminopeptidase I (API), a vacuolar hydrolase, is selectively transported to the vacuole via the autophagosome. API forms a cytosol to vacuole targeting (Cvt) complex in the cytoplasm. The complex is engulfed by the autophagosome under starvation conditions. In this study, the Cvt complex is visualized as a dot in the cytoplasm by fluorescence microscopy with API-GFP. The Cvt complex associates with the pre-autophagosomal structure (PAS), which plays a central role in autophagosome formation. In a cvt19 mutant, which is specifically defective in API transport, but not in autophagy, the Cvt complex forms normally but never associates with the PAS. This indicates that Cvt19p mediates association between the Cvt complex and the PAS.
4. Studies of mammalian autophagyWe cloned mouse Apg10 and showed that the Apg12 conjugation system is completely conserved between yeast and mammals. In addition, we also cloned a novel Apg5-interacting protein, Apg16L. Although it was considered to be a functional counterpart of yeast Apg16, Apg16L has a large C-terminal region containing a WD repeat domain that is absent from yeast Apg16. This unique feature of Apg16L will provide new insights into role of Apg12-Apg5 in autophagosome formation. Once an autophagosome is formed, it matures by fusion with endosomes. Using a dominat negative mutant of mouse SKD1, an AAA ATPase involved in the sorting and transport from endosomes, we showed that SKD1-dependent endosomal membrane trafficking is required for further fusion with lysosome. To monitor autophagy in vivo, we generated transgenic mice expressing a fluorescent marker for autophagosome. It would be a useful tool for studies on mammalian autophagy. 5. Leaf senescence is accelerated by the disruption of an Arabidopsis autophagy gene.The molecular machinery responsible for yeast and mammalian autophagy has recently begun to be elucidated at the cellular level, but the role that autophagy plays at the organismal level has yet to be determined. In this study, a genome-wide search revealed significant conservation between yeast and plant autophagy genes. Twenty-five plant genes that are homologous to 12 yeast genes essential for autophagy were discovered. We identified an Arabidopsis mutant carrying a T-DNA insertion within AtAPG9, which is the only ortholog of yeast Apg9 in Arabidopsis (atapg9-1). AtAPG9 is transcribed in every wild-type organ tested but not in the atapg9-1 mutant. Under nitrogen or carbon-starvation conditions, chlorosis was observed earlier in atapg9-1 cotyledons and rosette leaves compared with wild-type plants. Furthermore, atapg9-1 exhibited a reduction in seed set when nitrogen starved. Even under nutrient growth conditions, bolting and natural leaf senescence were accelerated in atapg9-1 plants. Senescence-associated genes SEN1 and YSL4 were up-regulated in atapg9-1 before induction of senescence, unlike in wild type. All of these phenotypes were complemented by the expression of wild-type AtAPG9 in atapg9-1 plants. These results imply that autophagy is required for maintenance of the cellular viability under nutrient-limited conditions and for efficient nutrient use as a whole plant.
Original papers :Hamasaki M, Noda T, Ohsumi Y. (2003) The early seretory pathway contributes to autophagy in yeast. Cell Struct. Funct. (in press) Hanaoka H, Noda T, Shirano Y, Kato T, Hayashi H, Shibata D, Tabata S, Ohsumi Y. (2002) Leaf senescence and starvation-induced chlorosis are accelerated by the disruption of an Arabidopsis autophagy gene. Plant Physiol 129, 1181-1193. Kuma A, Mizushima N, Ishihara N, Ohsumi Y. (2002) Formation of the approximately 350-kDa Apg12-Apg5.Apg16 multimeric complex, mediated by Apg16 oligomerization, is essential for autophagy in yeast. J Biol Chem 277, 18619-18625. Mizushima N, Yoshimori T, Ohsumi Y. (2002) Mouse Apg10 as an Apg12-conjugating enzyme: analysis by the conjugation-mediated yeast two-hybrid method. FEBS Lett. 532, 450-454. Mimura T., Reid R.J., Ohsumi Y., Smith F.A. (2002) Induction of the Na+/Pi co-transport system in the plasma membrane of Chara corallina requires external Na+ and low levels of Pi. Plant Cell & Environment 25:1475-1481. Nara A, Mizushima N, Yamamoto A, Kabeya Y, Ohsumi Y, Yoshimori T. (2002) SKD1 AAA ATPase-dependent endosomal transport is involved in autolysosome formation. Cell Struct Funct. 27, 29-37. Suzuki K, Kamada Y, Ohsumi Y. (2002) Studies of cargo delivery to the vacuole mediated by autophagosomes in Saccharomyces cerevisiae. DevCell 3, 815-824. Suzuki T, Nakagawa M, Yoshikawa A, Sasagawa N, Yoshimori T, Ohsumi Y, Nishino I, Ishiura S, Nonaka I. (2002) The first molecular evidence that autophagy relates rimmed vacuole formation in chloroquine myopathy J Biochem (Tokyo). 131, 647-651. Reviews:Kamada Y. Sekito T. Ohsumi Y. (2003)Autophgy in yeast- a Tor-mediated response to nutrient starvation- Curr. Top. Microbiol. Immunol. (in press) Mizushima N, Yoshimori T, Ohsumi Y. (2003) Role of the Apg12 conjugation system in mammalian autophagy Int. J. Biochem. Cell Biol.(in press) Mizushima N, Ohsumi Y, Yoshimori T. (2002) Autophagosome formation in mammalian cells Cell Struct Funct in press Noda T, Suzuki K, Ohsumi Y. (2002) Yeast autophagosomes: de novo formation of a membrane structure.Trends Cell Biol.12, 231-235 Ohsumi, Y. (2003) Cellular Recycling System- Molecular Mechanism of Autophagy-.In Cell Growth Cold Spring Harbour Press. (in press) |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Copyright(C) National Institute for Basic Biology (webmaster@nibb.ac.jp) |