DIVISION OF CELL MECHANISMS
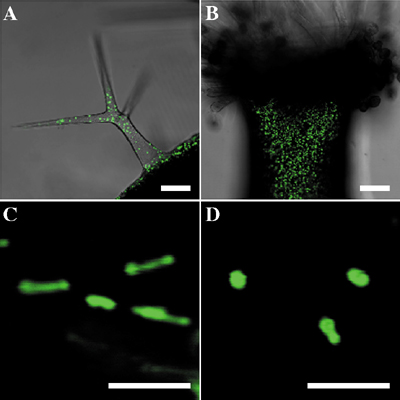
Higher plant cells contain several distinct organelles that play vital roles in cellular physiology. During proliferation and differentiation of the cells, the organelles often undergo dynamic changes. The biogenesis of new organelles may occur, existing organelles may undergo a transformation of function, while other organelles may degenerate. Because the dynamic transformation of organellar function (differentiation of organelles) is responsible for flexibility of differentiation events in higher plant cells, the elucidation of regulatory mechanisms underlying organelle transformation are currently studied in this division. I. Reversible transformation of plant peroxisomesDramatic metabolic changes which underlie the shift from heterotrophic to autotrophic growth occur in greening of seed germination. Accompanying these metabolic changes, many constitutive organelles are functionally transformed. Etioplasts differentiate into chloroplasts and mitochondria acquire the ability to oxidize glycine. Glyoxysomes which are microbodies engaged in the degradation of reserve oil via b-oxidation and the glyoxylate cycle, are transformed into leaf peroxisomes that function in several crucial steps of photorespiration. After the functional transition of glyoxysomes to leaf peroxisomes during the greening of pumpkin cotyledons, the reverse transition of leaf peroxisomes to glyoxysomes occurs during senescence. The functional transformation between glyoxysomes and leaf peroxisomes is controlled by gene expression, alternative splicing, protein translocation and protein degradation. We now engage in proteomic and transcriptomic analyses of the reversible peroxisomal transition in Arabidopsis cotyledons. II. Intracellular transport of peroxisomal proteinsEnzymes localized in plant peroxisomes are synthesized in the cytosol, and function after their post-translational transport into peroxisomes. Almost all of the peroxisomal matrix proteins are known to contain one of two targeting signals (PTS1 and PTS2) within the molecules. PTS1 is a unique tripeptide sequence found in carboxyl terminus of the mature proteins. The permissible combinations of amino acids for PTS1 in plant cells are [C/A/S/P]-[K/R]-[I/L/M]. In contrast, PTS2 is involved in a cleavable amino terminal presequence of peroxisomal proteins that are synthesized as precursor protein with larger molecular mass. PTS2 consists of a consensus sequence [R]-[L/Q/I]-X5-[H]-[L]. PTS1 and PTS2 are initially recognized by binding with their cytosolic receptors, AtPex5p and AtPex7p, respectively. Since AtPex5p could also bind with AtPex7p, they might form a receptor-cargo complex. Catalase is a peroxisomal protein that have been know to contain no obvious PTS1/PTS2. We recently found that 480QKL of the enzyme function as an alternative peroxisomal targeting signal. Yeast two-hybrid analysis revealed that the signal also binds with AtPex5p. The result indicated that catalase is also imported into peroxisomes by PTS1-dependent manner. Arabidopsis ped2 mutant is one of the mutants that show resistance to 2,4-dichlorophenoxybutyric acid. PED2 gene, responsible for this deficiency, encodes a peroxisomal membrane protein, AtPex14p. Mutation in the PED2 gene results in pleiotropic defects on fatty acid degradation, photorespiration, and the morphology of all kinds of peroxisomes by disrupting both PTS1- and PTS2-dependent peroxisomal protein import pathways. Binding assay revealed that AtPex14p directly binds with AtPex5p, but not with AtPex7p. AtPex14p controls both PTS1- and PTS2-dependent peroxisomal protein import pathways by binding with the receptor-cargo complex involving AtPex5p. These data suggest that the AtPex14p has a common role in maintaining physiological functions of all kinds of plant peroxisomes by determining peroxisomal protein targeting. III. Dynamic morphology and actin dependent movement of plant peroxisomesTo characterize peroxisomes in vivo, we visualized them in cells of various tissues of Arabidopsis by green fluorescent protein (GFP) through the addition of the peroxisomal targeting signal 1 (PTS1) or PTS2. The observation using confocal laser scanning microscopy revealed that the GFP fluorescence signals were detected as spherical spots in all cells of two kinds of transgenic plants. Interestingly, however, abnormal large peroxisomes are present as clusters at the onset of germination, and these clusters disappear in a few days. Moreover, tubular peroxisomes which looked like the dumbbell, as if two peroxisomes were connected with the tubule, were also observed in the hypocotyl. Peroxisomes turned to be motile organelles whose movement might be caused by cytoplasmic flow. The movement of peroxisomes was more prominent in root cells than that in leaves, and divided into two categories: a relatively slow, random, vibrational movement and a rapid movement. Treatment with anti-actin and anti-tubulin drugs revealed that actin filaments involve in the rapid movement of peroxisomes. These findings indicate that peroxisomes undergo dramatic changes in size, shape, number and position within the cell.
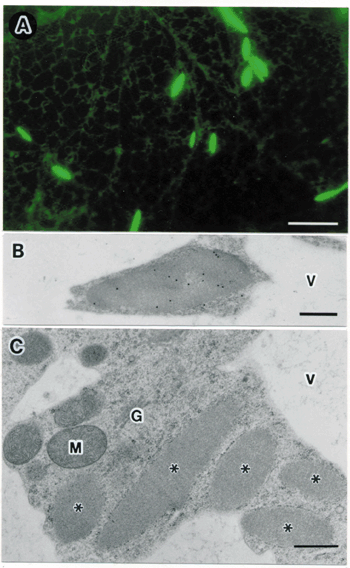
IV. ER derived organelles for transport of proteins to vacuoles.A novel vesicle, referred to as a precursor-accumulating (PAC) vesicle, mediates the transport of storage protein precursors from endoplasmic reticulum (ER) to protein storage vacuoles in maturing pumpkin seeds. PAC vesicles had diameters of 300 to 400 nm, are derived from ER and contained an electron-dense core of storage proteins. PV72, a type I integral membrane protein with three repeats of epidermal growth factor (EGF)-like motifs, was found on the membrane of the PAC vesicles. We expressed modified PV72 and analyzed their ability to bind to the internal propeptide of pro2S albumin (2S-I), a storage protein precursor, by affinity chromatography and surface plasmon resonance. The recombinant PV72 specifically bound to the 2S-I peptide with a KD value of 0.2 mM, which was low enough for it to function as a receptor. The EGF-like motifs modulate the Ca2+-dependent conformational change of PV72 to ER bodies are one of ER-derived compartment specific to the Brassicaceae, including Arabidopsis. ER bodies are rod-shaped structures that are characteristically 5 mm long and 0.5 mm wide. Electron microscopic studies revealed that ER bodies have a fibrous pattern inside and are surrounded by ribosomes (Fig. 2). Fluorescent imaging of ER bodies were observed in transgenic plants of Arabidopsis (GFP-h) expressing green fluorescent protein fused with an ER retention signal (GFP-HDEL). ER bodies were widely distributed in the epidermal cells of whole seedlings. In contrast, rosette leaves had no ER bodies. We found that wound stress induced the formation of ER bodies in rosette leaves. ER bodies were also induced treatment of methyl jasmonate (MeJA), a plant hormone involved in the defense response against wounding. The induction of ER bodies by MeJa was suppressed by ethylene. ER bodies contained precursors of two vacuolar proteinases, RD21 and VPE. During salt-induced cell death, ER bodies fused with each other and with lytic vacuoles, thereby mediating the delivery of the proteinase precursors directly into the vacuoles. We isolated an Arabidopsis mutant, named nai1, in which ER bodies were hardly detected in whole plants. ER bodies were concentrated in a 1000 x g pellet (P1) after subcellular fractionation. PYK10, a b-glucosidase with an ER-retention signal (KDEL), was specifically accumulated in the P1 fraction of GFP-h plants, but not in the P1 fraction of nai1 plants. Compare with the accumulation of GFP-HDEL, which was associated with both cisternal ER and ER bodies, the accumulation of PYK10 was much more specific to ER bodies. These findings indicate that PYK10 is the main component of ER bodies. Taken together, these results suggest that the formation of ER bodies is a novel and unique type of endomembrane system in the responses of certain cells to environmental stress.
IV. Role of molecular chaperones in organelle differentiation.Molecular chaperones are cellular proteins that function in the folding and assembly of certain other polypeptides into oligomeric structures but that are not, themselves, components of the final oligomeric structure. To clarify the roles of molecular chaperones on organelle differentiation, we have purified and characterized chaperonin and Hsp70s and analyzed their roles in the translocation of proteins into chloroplasts. Previously, we characterized a mitochondrial co-chaperonin (Cpn10) and chloroplast co-chaperonins (Cpn20) and (Cpn10) from Arabidopsis. In 2002, we started to characterize molecular chaperones in peroxisomes with collaboration with Dr. A. Kato at Niigata Univ. We found that a novel small heat shock protein (sHsp) localizes in peroxisomes. Molecular and functional characterization of the sHsp is now under experiments. Publication List:Fukao, Y., Hayashi, M. and Nishimura, M. (2002) Proteomic analysis of leaf peroxisomes in greening cotyledons of Arabidopsis thaliana. Plant Cell Physiol. 43, 689-696. Hayashi, M., Nito, K., Takei-Hoshi, R., Yagi, M., Kondo, M., Suenaga, A., Yamaya, T. and Nishimura, M. (2002) Ped 3p is a peroxisomal ATP-binding cassete transporter that might supply substrates for fatty acid b-oxidation. Plant Cell Physiol. 43, 1-11. Hayashi, H., De Bellis L., Kato, A., Hayashi, Y., Nito, K., Hayashi, M., Hara-Nishimura, I. and Nishimura, M. (2002) Molecular characterization of an Arabidopsis acyl CoA synthetase localized on glyoxysomal membranes. Plant Physiol. 130, 2019-2026. Kamigaki, A., Mano, S., Terauchi, K., Nishi, Y., Tachibe-Kinoshita, Y., Kondo, M., Nito, K., Hayashi, M., Nishimura, M. and Esaka, M. (2003) Identification of peroxisomal targeting signal of pumpkin catalase and the binding analysis with PTS1 receptor. Plant J. 33. 161-175. Kimura, Y., Matsuno, S., Tsurusaki, S., Kimura, M., Hara-Nishimura, I. and Nishimura, M. (2002) Subcellular localization of endo-b-N-acetylglucosaminidase and high-mannose type free N-glycans in plant cell. Biochim. Biophys. Acta 1570, 38-46. Kuroyanagi, M., Nishimura, M. and Hara-Nishimura, I. (2002) Activation of Arabidopsis vacuolar processing enzyme by self-catalytic removal of an auto-inhibitory domain of the C-terminal propeptide. Plant Cell Physiol. 43, 143-151. Mano, S., Nakamori, C., Hayashi, M., Kato, A., Kondo, M. and Nishimura, M. (2002) Distribution and characterization of peroxisomes in Arabidopsis by visualization with GFP: Dynamic morphology and actin dependent movement. Plant Cell Physiol. 43, 331-341. Maser, P., Eckerman, B., Vaidyanathan, R., Fairbairn, D. J., Kubo, M., Yamagami, M., Yamaguchi, K., Nishimura, M., Uozumi, N., Robertson, W., Sussman, M. and Schroeder, J. I. (2002) Altered shoot/root Na+ distribution and bifurcating salt sensitivity in Arabidopsis by genetic disruption of the Na+ transportor AtHKT1. FEBS Lett. 531, 157-161. Matsushima, R., Hayashi, Y., Kondo, M., Shimada, T., Nishimura, M. and Hara-Nishimura, I. (2002) An ER-derived structure that is induced under stress conditions in Arabidopsis. Plant Physiol. 130, 1807-1814. Matsushima, R., Kondo, M., Nishimura, M. and Hara-Nishimura, I. (2002) A novel ER-derived compartment, the ER body, selectively accumulates a b-glucosidase with an ER retention signal in Arabidopsis. Plant J. in press Nito, K., Hayashi, M. and Nishimura, M. (2002) Direct interaction and determination of binding domains among peroxisomal import factors in Arabidopsis thaliana. Plant Cell Physiol. 43, 355-366. Ono, K., Kondo M., Osafune, T., Miyatake, K., Inui, H., Kitaoka, S., Nishimura, M. and Nakano, Y. (2002) Presence of Glyoxylate Cycle Enzymes in the Mitochondria of Euglena gracilis. J. Eukaryo. Microbiol.in press Shimada, T., Watanabe, E., Tamura, K., Hayashi, Y., Nishimura, M., and Hara-Nishimura, I. (2002) A vacuolar-sorting receptor on the membrane of the PAC vesicles that accumulate precursors of seed storage proteins. Plant Cell Physiol. 43, 1086-1095. Watanabe, E., Shimada, T., Kuroyanagi, M., Nishimura, M. and Hara-Nishimura, I. (2002) Calcium-mediated association of a putative vacuolar sorting receptor PV72 with a propeptide of 2S albumin. J. Biol. Chem. 277, 8708-8715. |
|
Copyright(C) National Institute for Basic Biology (webmaster@nibb.ac.jp) |