CENTER FOR RADIOISOTOPE FACILITIES (CRF)
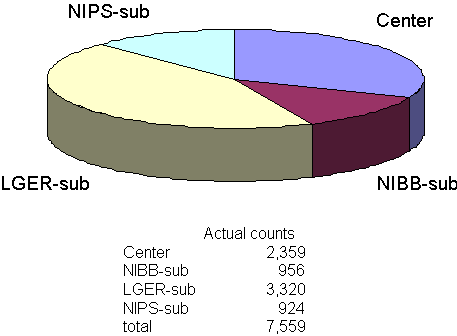
I. Research supporting activityTechnical and supporting staffs of the CRF are serving the purchase of radioisotopes from JRA (Japan Radioisotope Association) and the transfer of radioisotope wastes to JRA. The physical maintenance of the controlled areas where radioisotopes are used is also one of our business. The CRF consists of four controlled areas: Center, NIBB-sub, LGER (Laboratory of Gene Expression and Regulation)-sub, and NIPS (National Institute for Physiological Science)-sub. Users going in and out the controlled areas counted by the monitoring system are 7,559 in 2001. The items in each controlled area is presented in Figure 1.
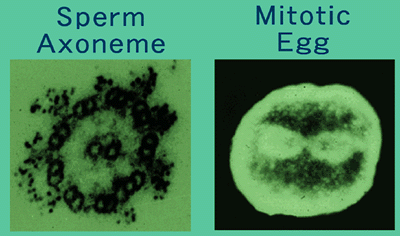
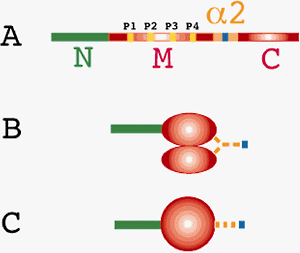
II. Academic activityAcademic activity by teaching staff is focused on the analysis of the structure and function of a dynein motor protein. Dyneins are a group of microtubule-activated ATPases that serve to convert chemical energy into mechanical energy and divided into axonemal and cytoplasmic dyneins. Figure 2 shows the localization of two isoforms of dynein in the outer arms of sperm axonemes (Ogawa et al., 1977) and the mitotic apparatus of cleaving egg (Mohri et al., 1976) visualized by anti-axonemal dynein (Fragement A) antibodies. The native dyneins are very large and range in molecular mass up to 1 to 2 mega Da. They are complex proteins containing heavy, intermediate, and light chains defined by the molecular mass. Our prensent project is the molecular cloning of polypeptides contained in outer arm dynein of sea urchin sperm flagella to understand the mechanism how dynein interacts with microtubules, resulting in producing the force. Outer arm dynein consists of two heavy chains with ATPase activity. The motor activity is closely related to this polypeptide. The first successful molecular cloning of this huge polypeptide (520 kDa) was performed in our laboratory in 1991. Since then cDNA clones for axonemal and cytoplasmic dyneins have been isolated in a variety of organisms. The sequences of heavy chains, without exception, contain four P-loop motives referred to as ATP-binding sites in the midregion of the molecules. Figure 3A and B draw the structure of heavy chain deduced from the amino acid sequence (Ogawa, 1992). Taking the recent works by Koonce et al. (1998) and Vallee et al. (1998) into consideration, this model might be seen as depicted in Figure 3C. In particular, Vallee et al. (1998) have described the importance of a hairpin structure formed between M and C domains which binds to microtubules and presented a novel mechanism for dynein force production different from that of myosin and kinesin. Outer arm dynein contains three intermediate chains (IC1, IC2, and IC3) that range in molecular mass from 70 to 120 kDa. IC2 and IC3 were cloned by Ogawa et al. (1995) and contain the WD repeats in the carboxy-terminal halves of the molecules. By contrast, IC1 is not a member of the WD family. IC1 has a unique sequence such that the N-terminal part is homologous to the sequence of thioredoxin, the middle part consists of three repetitive sequences homologous to the sequence of NDP kinase, and the C-terminal part contains a high proportion of negatively charged glutamic acid residues (Ogawa et al., 1996). Thus, IC1 is a novel dynein intermediate chain distinct from IC2 and IC3 and may be a multifunctional protein. Six light chains with molecular masses of 23.2, 20.8, 12.3, 11.5, 10.4, and 9.3 kDa are associating with outer arm dynein. We have already isolated cDNA clone of five LCs. LC1 (23.2 kDa) and LC3 (12.3 kDa) are highly homologous to mouse Tctex2 and Tctex1, respectively. These mouse proteins are encoded by the t complex region that is involved in transmission ratio distortion (TRD), male sterility and the development of germ cells. Our finding raises the possibility that axonemal dynein proteins are involved in this phenomenon. TRD may be caused by the dysfunction of multiple axonemal dynein proteins.
|
|
Copyright(C) National Institute for Basic Biology (webmaster@nibb.ac.jp) |